8 Rotifera, Platyhelminthes, Molluska and Annelida
8.1 Rotifers
The rotifers (Rotifera, commonly called wheel animals) make up a phylum of microscopic and near-microscopic pseudocoelomate animals. The word “rotifer” is derived from a Latin word meaning “wheel-bearer”, due to the corona around the mouth that in concerted sequential motion resembles a wheel (though the organ does not actually rotate). They were first described by Rev. John Harris in 1696, and other forms were described by Antonie van Leeuwenhoek in 1703. Most rotifers are around 0.1–0.5 mm long (although their size can range from 50 μm to over 2 mm), and are common in freshwater environments throughout the world with a few saltwater species; for example, those of genus Synchaeta. Some rotifers are free swimming and truly planktonic, others move by inchworming along a substrate, and some are sessile, living inside tubes or gelatinous holdfasts that are attached to a substrate. About 25 species are colonial (e.g., Sinantherina semibullata), either sessile or planktonic. Rotifers are an important part of the freshwater zooplankton, being a major foodsource and with many species also contributing to the decomposition of soil organic matter.
Rotifers have bilateral symmetry and a variety of different shapes. The body of a rotifer is divided into a head, trunk, and foot, and is typically somewhat cylindrical. There is a well-developed cuticle, which may be thick and rigid, giving the animal a box-like shape, or flexible, giving the animal a worm-like shape; such rotifers are respectively called loricate and illoricate. Rigid cuticles are often composed of multiple plates, and may bear spines, ridges, or other ornamentation. Their cuticle is non-chitinous and is formed from sclerotized proteins.
The most distinctive feature of rotifers is the presence of a ciliated structure, called the corona, on the head. In the more primitive species, this forms a simple ring of cilia around the mouth from which an additional band of cilia stretches over the back of the head. In the great majority of rotifers, however, this has evolved into a more complex structure. The trunk forms the major part of the body, and encloses most of the internal organs. The foot projects from the rear of the trunk, and is usually much narrower, giving the appearance of a tail. The cuticle over the foot often forms rings, making it appear segmented, although the internal structure is uniform. Many rotifers can retract the foot partially or wholly into the trunk. The foot ends in from one to four toes, which, in sessile and crawling species, contain adhesive glands to attach the animal to the substratum. In many free-swimming species, the foot as a whole is reduced in size, and may even be absent.
Rotifers are dioecious and reproduce sexually or parthenogenetically. They are sexually dimorphic, with the females always being larger than the males. In some species, this is relatively mild, but in others the female may be up to ten times the size of the male. In parthenogenetic species, males may be present only at certain times of the year, or absent altogether. The female reproductive system consists of one or two ovaries, each with a vitellarium gland that supplies the eggs with yolk. Together, each ovary and vitellarium form a single syncitial structure in the anterior part of the animal, opening through an oviduct into the cloaca. Males do not usually have a functional digestive system, and are therefore short-lived, often being sexually fertile at birth. They have a single testicle and sperm duct, associated with a pair of glandular structures referred to as prostates (unrelated to the vertebrate prostate). The sperm duct opens into a gonopore at the posterior end of the animal, which is usually modified to form a penis. The gonopore is homologous to the cloaca of females, but in most species has no connection to the vestigial digestive system, which lacks an anus. Fertilization is internal. The male either inserts his penis into the female’s cloaca or uses it to penetrate her skin, injecting the sperm into the body cavity. The egg secretes a shell, and is attached either to the substratum, nearby plants, or the female’s own body. A few species, such as Rotaria, are ovoviviparous, retaining the eggs inside their body until they hatch. Most species hatch as miniature versions of the adult. Sessile species, however, are born as free-swimming larvae, which closely resemble the adults of related free-swimming species. Females grow rapidly, reaching their adult size within a few days, while males typically do not grow in size at all.
8.2 Platyhelminthes
The flatworms, flat worms, Platyhelminthes, Plathelminthes, or platyhelminths (from the Greek platy, meaning “flat” and helminth-, meaning “worm”) are a phylum of relatively simple bilaterian, unsegmented, soft-bodied invertebrates. Unlike other bilaterians, they are acoelomates (having no body cavity), and have no specialized circulatory and respiratory organs, which restricts them to having flattened shapes that allow oxygen and nutrients to pass through their bodies by diffusion. The digestive cavity has only one opening for both ingestion (intake of nutrients) and egestion (removal of undigested wastes); as a result, the food cannot be processed continuously.
Free-living flatworms are mostly predators, and live in water or in shaded, humid terrestrial environments, such as leaf litter. Cestodes (tapeworms) and trematodes (flukes) have complex life-cycles, with mature stages that live as parasites in the digestive systems of fish or land vertebrates, and intermediate stages that infest secondary hosts. The eggs of trematodes are excreted from their main hosts, whereas adult cestodes generate vast numbers of hermaphroditic, segment-like proglottids that detach when mature, are excreted, and then release eggs. Unlike the other parasitic groups, the monogeneans are external parasites infesting aquatic animals, and their larvae metamorphose into the adult form after attaching to a suitable host.
Over half of all known flatworm species are parasitic, and some do enormous harm to humans and their livestock. Schistosomiasis, caused by one genus of trematodes, is the second-most devastating of all human diseases caused by parasites, surpassed only by malaria. Neurocysticercosis, which arises when larvae of the pork tapeworm Taenia solium penetrate the central nervous system, is the major cause of acquired epilepsy worldwide. The threat of flatworm parasites to humans in developed countries is rising because of the popularity of raw or lightly cooked foods, and imports of food from high-risk areas. In less developed countries, people often cannot afford the fuel required to cook food thoroughly, and poorly designed water-supply and irrigation projects increase the dangers presented by poor sanitation and unhygienic farming.
In traditional medicinal texts, Platyhelminthes are divided into
- Turbellaria
- Cestoda
- Trematoda
- Monogenea
8.3 Turbellaria
Turbellaria have about 4,500 species, are mostly free-living, and range from 1 mm (0.039 in) to 600 mm in length. Most are predators or scavengers, and terrestrial species are mostly nocturnal and live in shaded, humid locations, such as leaf litter or rotting wood. However, some are symbiotes of other animals, such as crustaceans, and some are parasites. Free-living turbellarians are mostly black, brown or gray, but some larger ones are brightly colored.
8.4 Dugesia
Dugesia is a representatives of the class Turbellaria. These common flatworms are found in freshwater habitats of Africa, Europe, Middle East, Asia and Australia. Dugesia species have an elongated body with a slightly triangle-shaped head. Usually they have grey, brown or black colors on the dorsal body surface, the ventral surface uses to be paler. These animals have a couple of eyes constituted by a multicellular pigmented cup with many retinal cells to detect the amount of light in the nearby environment. Sometimes they present supernumerary eyes. At the anterior part of the body, behind the eyes level, they have two of structures called auricles that give the triangle look to the ‘head’ and that allow them to detect the intensity of water current. These auricles are free of pigment and rhabdites. Each side of the anterior margin of the head have between 5 and 10 shallow sensory fossae, their number depends on the species or the individual. The sensory fossae and the auricle grooves are supplied with many nerve endings.
Dugesia digestion tract consists of a central non-pigmented tubular pharynx. Like the other triclads Dugesia gut consists in three ramified branches. Each branch consists of ceca, which delivers the nutrients to the body. This worm has a sac digestive plan, that is, it does not have a separate opening for waste excretion.
In Dugesia the ovaries are ventrally situated, they start just behind the brain, usually at the level of the fourth intestinal branch. The bursal canal runs on the right side of the copulatory apparatus and above the atrium.
They are hermaphrodites. Many species can reproduce both sexually and asexually (by parthenogenesis or by fission).
8.5 View Prepared Slides of Dugesia
- Dugesia w.m. (Figure 8.1)
- Identify: highly branched gastrovascular cavity (GVC), pharynx, pharyngeal pouch, eyespots, auricles. Be sure to differentiate between anterior and posterior ends.
- Dugesia x.s. 3 regions (Figure 8.2)
- Identify: Anterior region: gastrovascular cavity (GVC) surrounded by endoderm; pharynx region: pharynx, GVC surrounded by endoderm, epidermis, testis, dorsoventral muscles; posterior end: epidermis, GVC surrounded by endoderm, dorsoventral muscles, ventral nerve cord
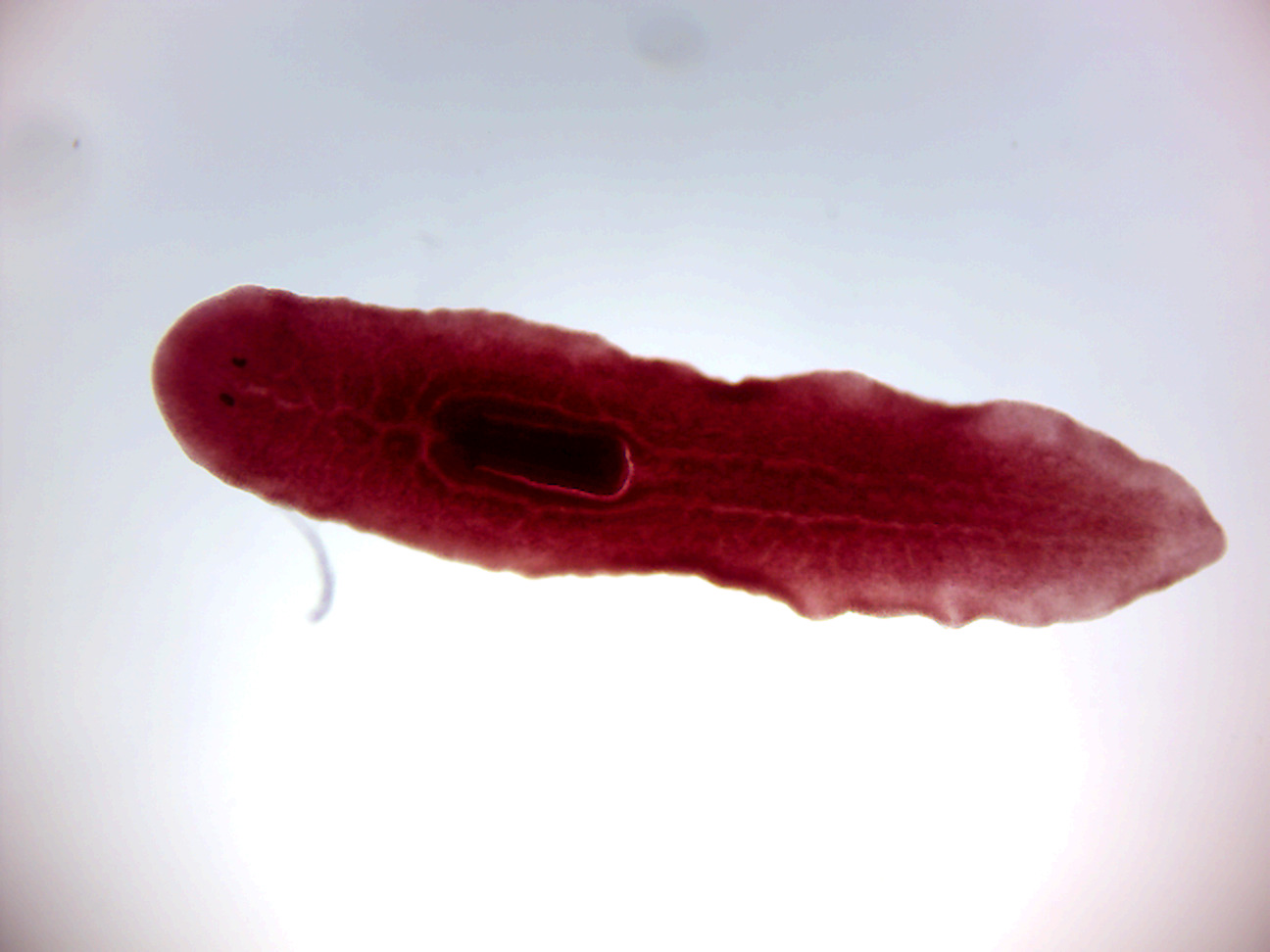
Figure 8.1: Dugesia.
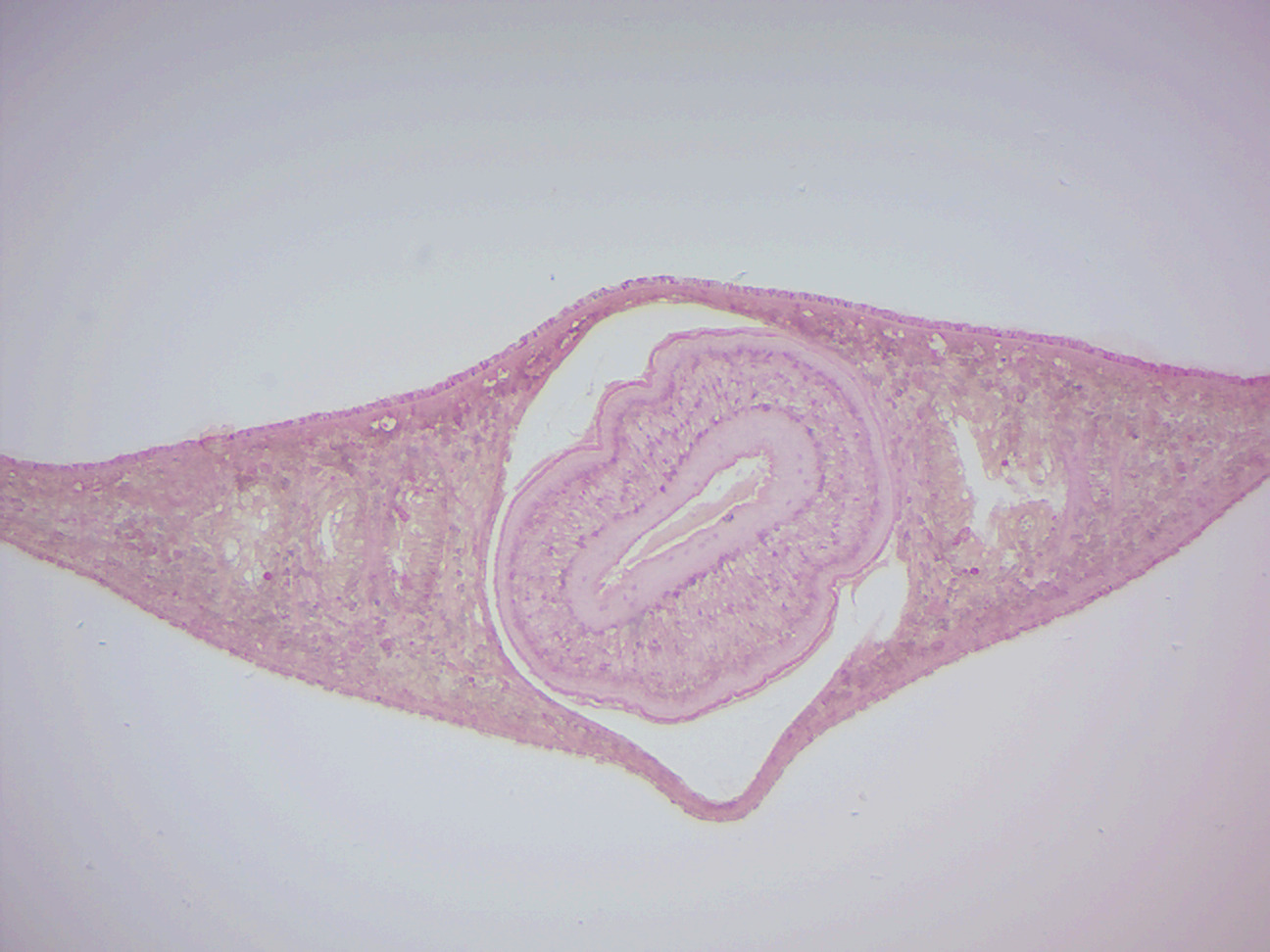
Figure 8.2: Dugesia cross section through pharynx region.
8.6 Cestoda
Cestoda are commonly called tapeworms. All cestodes are parasitic and their life histories vary, but typically they live in the digestive tracts of vertebrates as adults, and often in the bodies of other species of animals as juveniles. Over a thousand species have been described, and all vertebrate species may be parasitized by at least one species of tapeworm.
Humans are subject to infection by several species of tapeworms if they eat undercooked meat such as pork (Taenia solium), beef (T. saginata), and fish (Diphyllobothrium spp.), or if they live in, or eat food prepared in, conditions of poor hygiene (Hymenolepis or Echinococcus species).
T. saginata, the beef tapeworm, can grow up to 20 m. Species using small vertebrates as hosts, though, tend to be small.
Tapeworm parasites of vertebrates have a long history: recognizable clusters of cestode eggs, one with a developing larva, have been discovered in fossil feces (coprolites) of a shark dating to the mid- to late Permian, some 270 million years ago.
The worm’s head, known as a scolex, attaches to the intestine of the definitive host. In some species, the scolex is dominated by bothria, or “sucking grooves” that function like suction cups. Other species have hooks and suckers that aid in attachment.
The main nerve centre of a cestode is a cerebral ganglion in its scolex. Motor and sensory innervation depends on the number of nerves in and complexity of the scolex. Smaller nerves emanate from the ganglion to supply the general body muscular and sensory ending. The cirrus and vagina are innervated, and sensory endings around the genital pore are more plentiful than other areas. Sensory function includes both tactoreception (touch) and chemoreception (smell or taste). Some nerves are only temporary.
The body is composed of successive segments called proglottids. The sum of the proglottids is called a strobila, which is thin, and resembles a strip of tape. From this is derived the common name “tapeworm”. Proglottids are continually produced by the neck region of the scolex, as long as the scolex is attached and alive. Like some other flatworms, cestodes use flame cells (protonephridia), located in the proglottids, for excretion. Mature proglottids are released from the tapeworm’s end segment and leave the host in feces or migrate as independent motile proglottids. The proglottids farthest away from the scolex are the mature ones containing eggs. Mature proglottids are essentially bags of eggs, each of which is infective to the proper intermediate host.
Once anchored to the host’s intestinal wall, the tapeworm absorbs nutrients through its skin as the food being digested by the host flows over and around it. Soon, it begins to grow a tail composed of a series of segments, with each segment containing an independent digestive system and reproductive tract. Older segments are pushed toward the tip of the tail as new segments are produced by the neckpiece. By the time a segment has reached the end of the worm’s tail, only the reproductive tract is left. The segment then separates, carrying the tapeworm eggs out of the definitive host as what is basically a sack of eggs.
True tapeworms are exclusively hermaphrodites, with both male and female reproductive systems in their bodies. The reproductive system includes one or more testes, cirri, vas deferens, and seminal vesicles as male organs, and a single lobed or unlobed ovary with the connecting oviduct and uterus as female organs. The common external opening for both male and female reproductive systems is known as the genital pore, which is situated at the surface opening of the cup-shaped atrium. Though they are sexually hermaphroditic, self-fertilization is a rare phenomenon. To permit hybridization, cross-fertilization between two individuals is often practiced for reproduction. During copulation, the cirri of one individual connect with those of the other through the genital pore, and then spermatozoa are exchanged.
The lifecycle of tapeworms is simple in the sense that no asexual phases occur as in other flatworms, but complicated in that at least one intermediate host is required as well as the definitive host. Many tapeworms have a two-phase lifecycle with two types of hosts. The adult Taenia saginata lives in the gut of a primate such as a human, but more alarming is Taenia solium, which can form cysts in the human brain. Proglottids leave the body through the anus and fall onto the ground, where they may be eaten with grass by an animal such as a cow. If the tapeworm is compatible with the eating animal, this animal becomes an intermediate host. The juvenile form of the worm enters through the mouth, but then migrates and establishes as a cyst in the intermediate host’s body tissues such as muscles, rather than the gut. This can cause more damage to the intermediate host than it does to its definitive host. The parasite completes its lifecycle when the intermediate host passes on the parasite to the definitive host. This is usually done by the definitive host eating a suitably infected intermediate host, e.g., a human eating raw or undercooked meat.
8.7 View Prepared Slides Taenia
- Taenia pisoformis. Identify
- Taenia solium cysticercus w.m. (Figure 8.6).
- Identify: scolex, neck, bladder

Figure 8.3: Taenia scolex.

Figure 8.4: Taenia proglottid.
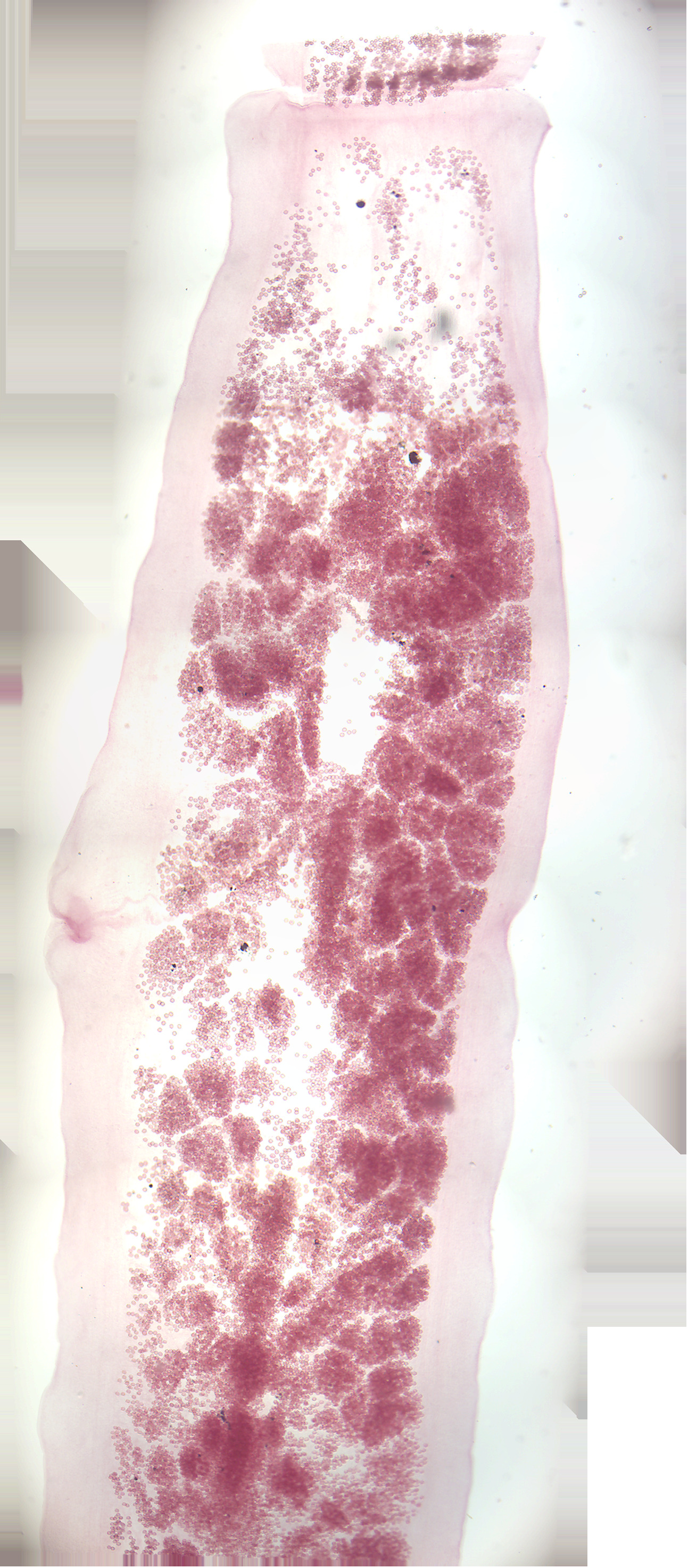
Figure 8.5: Taenia gravid proglottid.
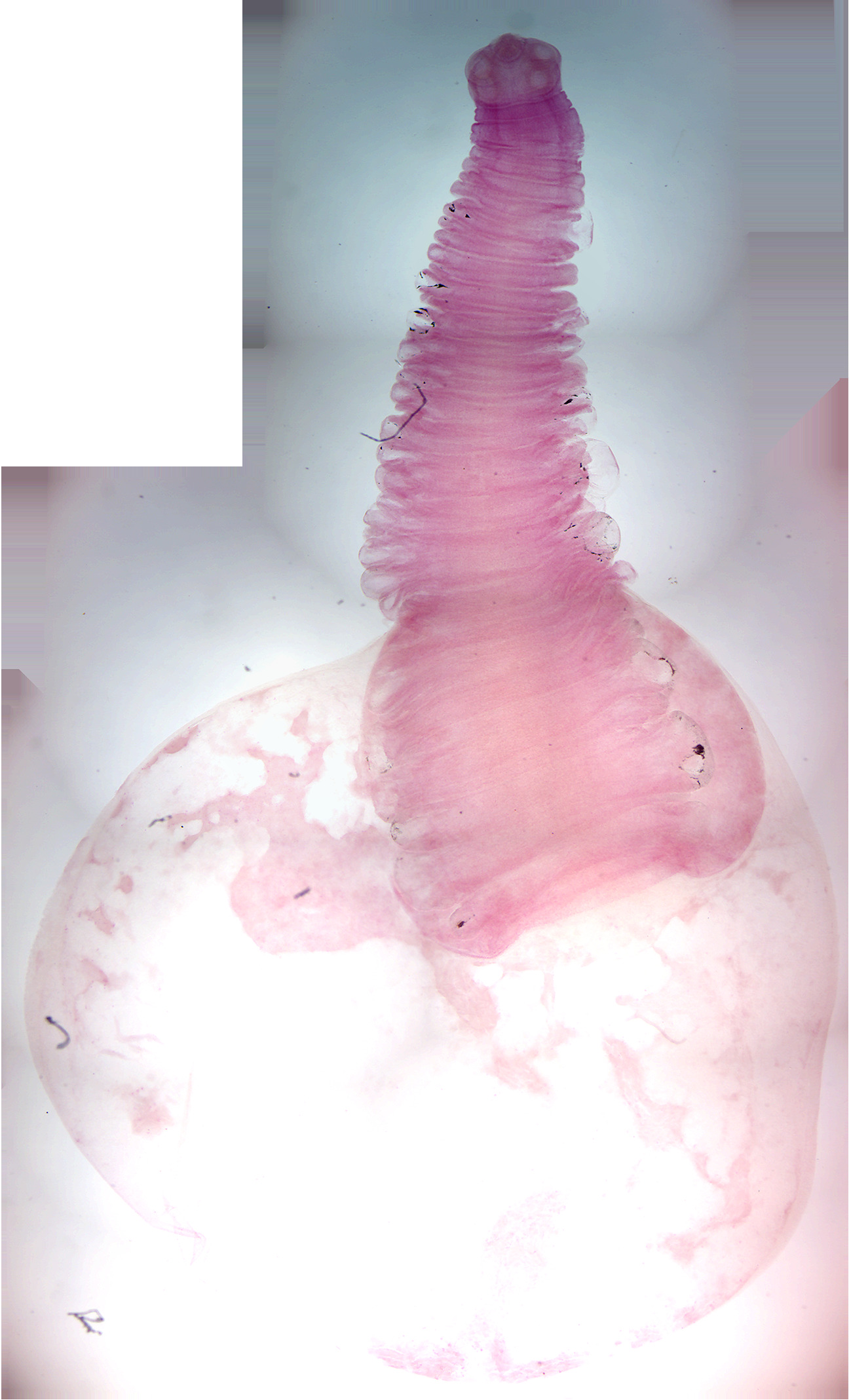
Figure 8.6: Taenia solium cysticercus.
8.8 Trematoda
The trematodes or flukes include 18,000 to 24,000 species, divided into two subclasses. Nearly all trematodes are parasites of mollusks and vertebrates. Most trematodes have a complex life cycle with at least two hosts. The primary host, where the flukes sexually reproduce, is a vertebrate. The intermediate host, in which asexual reproduction occurs, is usually a snail.
Trematodes are flattened oval or worm-like animals, usually no more than a few centimeters in length, although species as small as 1 millimeter) are known. Their most distinctive external feature is the presence of two suckers, one close to the mouth, and the other on the underside of the animal.
The body surface of trematodes comprises a tough syncitial tegument, which helps protect against digestive enzymes in those species that inhabit the gut of larger animals. It is also the surface of gas exchange; there are no respiratory organs. The mouth is located at the forward end of the animal, and opens into a muscular, pumping pharynx. The pharynx connects, via a short oesophagus, to one or two blind-ending caeca, which occupy most of the length of the body. In some species, the caeca are themselves branched. As in other flatworms, there is no anus, and waste material must be egested through the mouth.
Although the excretion of nitrogenous waste occurs mostly through the tegument, trematodes do possess an excretory system, which is instead mainly concerned with osmoregulation. This consists of two or more protonephridia, with those on each side of the body opening into a collecting duct. The two collecting ducts typically meet up at a single bladder, opening to the exterior through one or two pores near the posterior end of the animal.
The brain consists of a pair of ganglia in the head region, from which two or three pairs of nerve cords run down the length of the body. Trematodes generally lack any specialized sense organs, although some ectoparasitic species do possess one or two pairs of simple ocelli.
Most trematodes are simultaneous hermaphrodites, having both male and female organs. There are usually two testes, with sperm ducts that join together on the underside of the front half of the animal. This final part of the male system varies considerably in structure between species, but may include sperm storage sacs and accessory glands, in addition to the copulatory organ, which is either eversible, and termed a cirrus, or non-eversible, and termed a penis.
There is usually only a single ovary. Eggs pass from it into an oviduct. The distal part of the oviduct, called ootype, is dilated. The ootype is connected to an elongated uterus that opens to the exterior in the genital pore, close to the male opening. In most trematodes, sperm cells travel through the uterus to reach the ootype, where fertilization occurs.
Almost all trematodes infect mollusks as the first host in the life cycle, and most have a complex life cycle involving other hosts. Most trematodes are monoecious and alternately reproduce sexually and asexually. Schistosomes are dioecious.
In the definitive host, in which sexual reproduction occurs, eggs are commonly shed along with host feces. Eggs shed in water release free-swimming larval forms that are infective to the intermediate host, in which asexual reproduction occurs.
Human infections are most common in Asia, Africa, Latin and South America and the Middle East. However, trematodes can be found anywhere where untreated human waste is used as fertilizer. Schistosomiasis (also known as bilharzia, bilharziosis or snail fever) is an example of a parasitic disease caused by one of the species of trematodes (platyhelminth infection, or “flukes”), a parasitic worm of the genus Schistosoma. Clonorchis, Opisthorchis, Fasciola and Paragonimus species, the foodborne trematodes, are another. Other diseases are caused by members of the Choledocystus genus.
Clonorchis sinensis Clonorchis sinensis, the Chinese liver fluke, is a human liver fluke belonging to the class Trematoda, phylum Platyhelminthes. This parasite lives in the liver of humans, and is found mainly in the common bile duct and gall bladder, feeding on bile. These animals, which are believed to be the third most prevalent worm parasite in the world, are endemic to Japan, China, Taiwan, and Southeast Asia, currently infecting an estimated 30,000,000 humans. 85% of cases are found in China. The infection called clonorchiasis generally appears as jaundice, indigestion, biliary inflammation, bile duct obstruction, even liver cirrhosis, cholangiocarcinoma (CCA), and hepatic carcinoma.
An adult C. sinensis is a flattened (dorso-ventrally flat) and leaf-shaped fluke. The body is slightly elongated and slender, measuring 15–20 mm in length and 3–4 mm in width. It narrows down at the anterior region into a small opening called oral sucker, which act as the mouth. From the mouth run two tubes called caeca throughout the length of body. They are the digestive and excretory tracts. The posterior end is broad and blunt. A poorly developed ventral sucker lies behind the oral sucker, at about one-fourth of the body length form the anterior end. A common genital pore opens just in front of it. As a hermaphrodite, it has both male and female reproductive organs. A single rounded ovary is at the center of the body, and two testes are towards the posterior end. The uterus from the ovary, and seminal ducts from the testes meet and opens at the genital pore. The testes are highly branched. Another highly branched organs called vitellaria (or vitelline glands) are distributed on either side of the body.
The eggs are similar to those of other related flukes are often confused during diagnosis. They small and oval in shape, measuring about 30 x 15 μm in diameter. They are sharply curved and with a clear convex operculum towards the narrower end. At the broader end is a stem-shaped knob. The larva called miracidium can be seen inside the fertilised egg.
8.8.1 Life cycle
The eggs of a C. sinensis are released through the biliary tract, and excreted out along with the faeces. The eggs are embryonated and contain the larvae called miracidia. Unlike most other flukes in which the miracidia undergo development and swim in water to infect suitable host, the eggs of C. sinensis are simply deposited in water. The eggs are then eaten up by snails. Once inside of the snail body, the embryonic membrane is dissolved by the snail’s digestive enzymes so that the miracidium hatches from the egg. The ciliated miracidium can move about, penetrating the intestine, enters the haemocoel and digestive gland. Here it undergoes metamorphosis into a sporocyst. The sporocyst gives rise to small larvae called rediae. The rediae burst out from the sporocyst to become the next-stage larvae called cercaria. This system of asexual reproduction allows for an exponential multiplication of cercaria individuals from one miracidium. This aids the Clonorchis in reproduction, because it enables the miracidium to capitalize on one chance occasion of passively being eaten by a snail before the egg dies. The mature cercariae bore out of the snail body into the freshwater environment. However, they are non-feeding and must find a fish host within 2–3 days, otherwise they die. The ceraciae of C. sinensis are different from those of other flukes in that they do not swim. Instead, they initially hang upside-down in the water, and then sink to the bottom. They rise to the water surface to resume their initial position, and the movement is repeated again. They attack fish when they feel any disturbance in their life-style. When they detect fish, they attached themselves on the scales using their suckers. Boring their way into the fish’s body, they penetrate into the fish muscle within 6 to 13 minutes. Within an hour of penetration they develop hard coverings called cysts, and become metacercariae. This protective cyst is useful when the fish muscle is consumed. The metacercariae gradually develop and become infective to their next hosts after 3 to 4 weeks. The common second intermediate hosts are freshwater fish. The metacercariae are eaten along with raw or undercooked fish. The cysts of the metacercariae are gradually digested by the human gastric acids, and upon reaching the small intestines, the entire cyst is lost. The free metacercariae penetrate the intestinal mucosa and enters the bile ducts. It takes 1-2 day for migration into the bile ducts. They start feeding on the bile secreted from the liver, and gradually grow. They become adult in about a month, and start laying eggs. The average lifespan of an adult fluke is 30 years. An individual fluke can produce 4,000 eggs in a day. The definitive hosts are fish-eating mammals such as dogs, cats, rats, pigs, badgers, weasels, camels, buffaloes and humans.
8.9 View Prepared Slides of Trematoda
- Clonorchis sinensis w.m. (Figure 8.7).
- Identify: oral and ventral suckers, pharynx, esophagus, dead-end intestine, reproductive structures, excretory pore
- Clonorchis sinensis eggs (Figure 8.8).
- Identify: oral and ventral suckers, pharynx, esophagus, dead-end intestine, reproductive structures, excretory pore
- Fasciola hepatica w.m. (Figure 8.9).
- Identify: mouth, oral and ventral suckers, pharynx, uterus, intestine
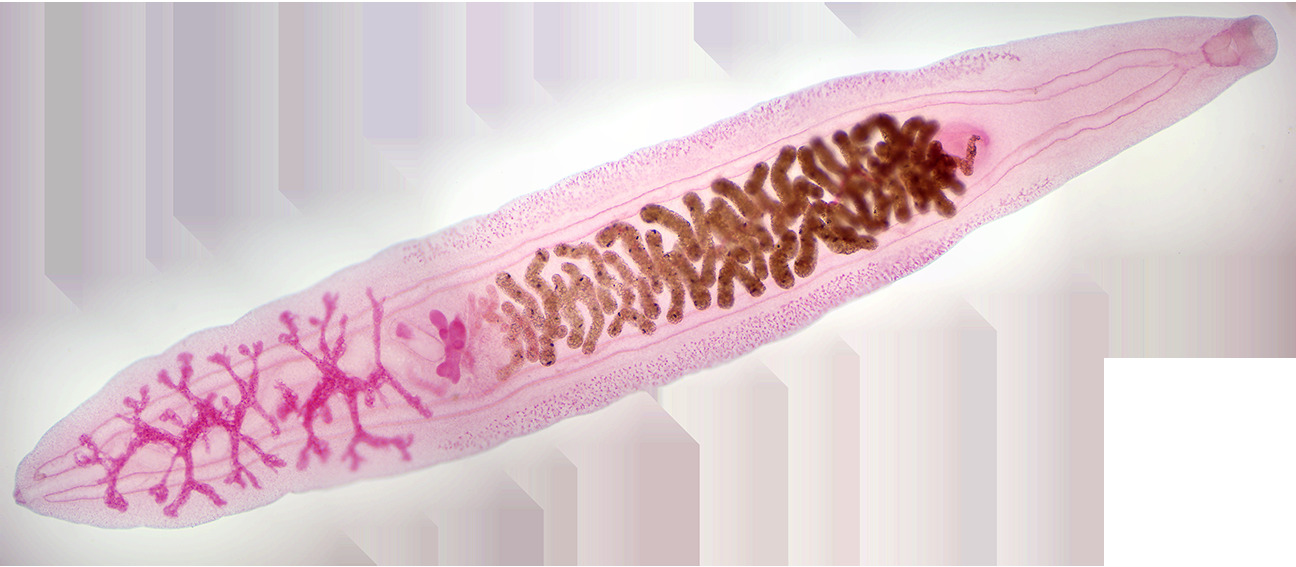
Figure 8.7: Clonorchis sinensis.
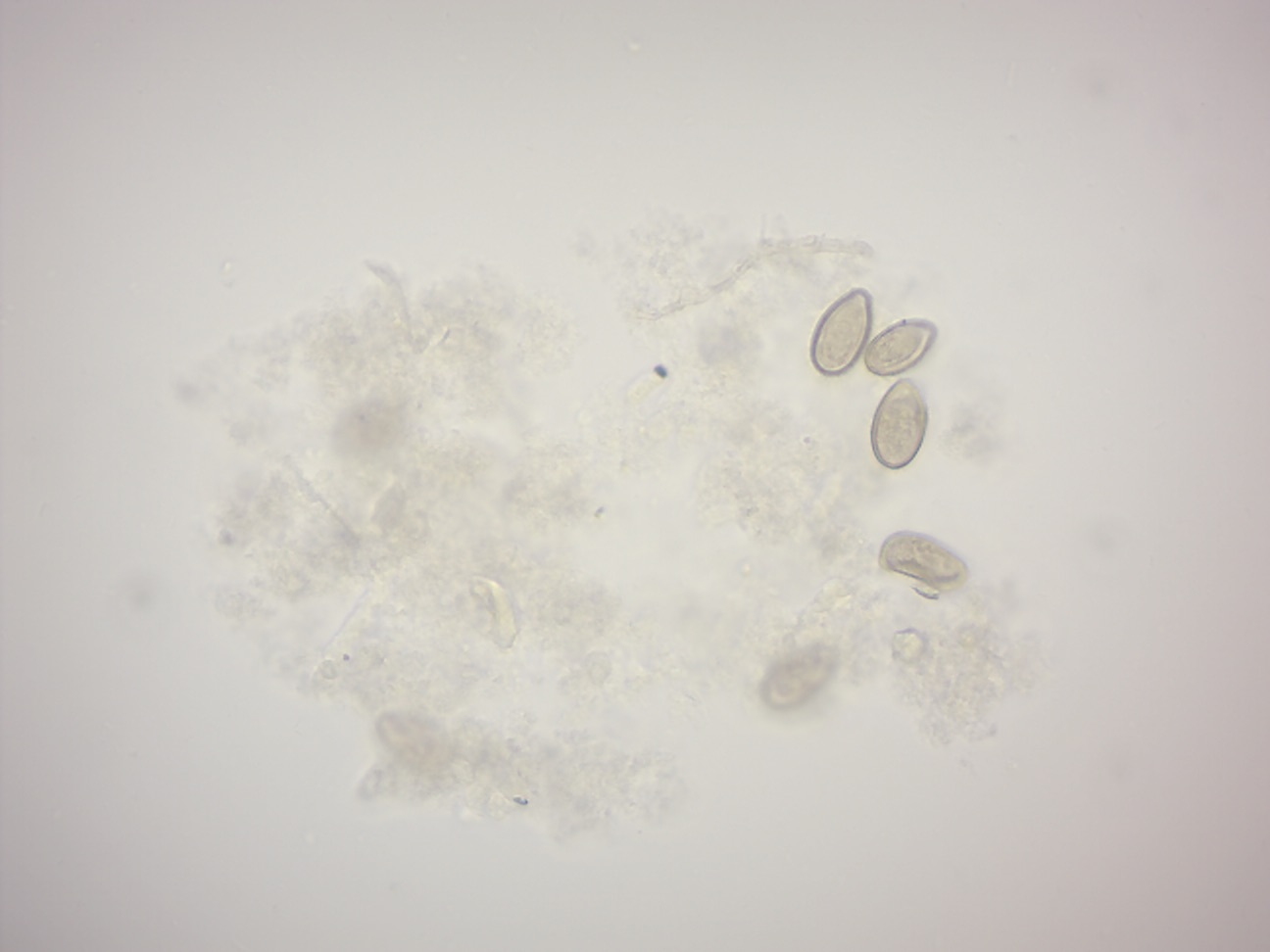
Figure 8.8: Clonorchis sinensis eggs.
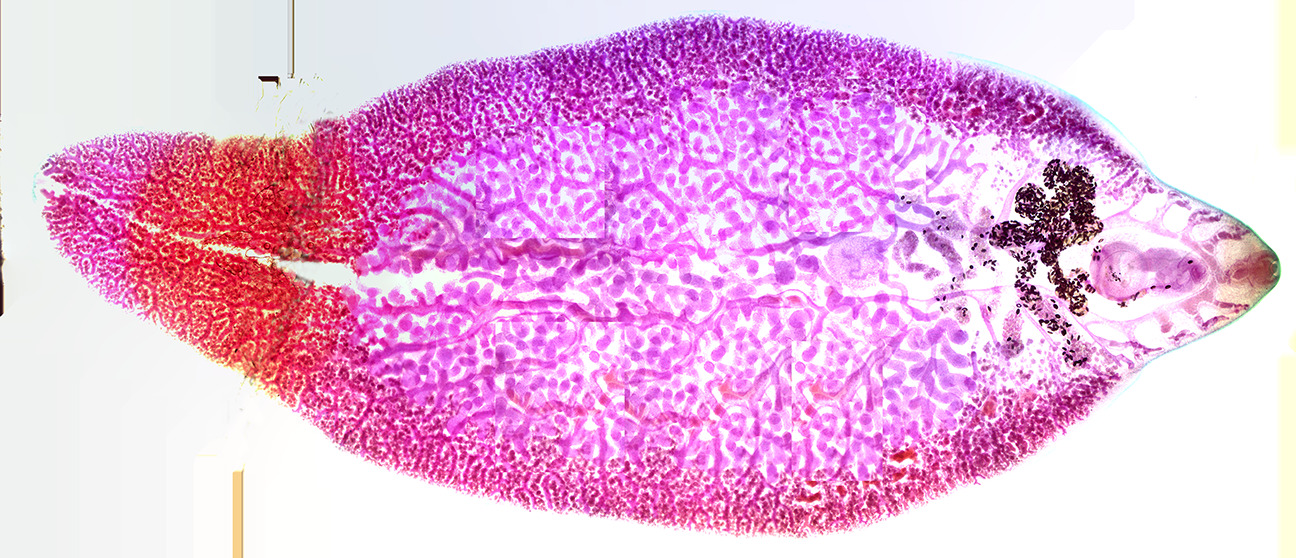
Figure 8.9: Fasciola hepatica.
8.10 Schistosoma
Schistosoma mansoni is a water-borne parasite of humans, and belongs to the group of blood flukes (Schistosoma). The adult lives in the blood vessels (mesenteric veins) near the human intestine. It causes intestinal schistosomiasis. Clinical symptoms are caused by the eggs. As the leading cause of schistosomiasis in the world, it is the most prevalent parasite in humans. It is classified as a neglected tropical disease. As of 2016, 206.5 million people have schistosomiasis and S. mansoni is the major parasite. It is found in Africa, the Middle East, the Caribbean, Brazil, Venezuela and Suriname.
Unlike other flukes (trematodes) in which sexes are not separate (monoecious), schistosomes are unique in that adults are divided into males and females, thus, (dioecious). However, the two adults live in permanent partnership, a condition called in copula; for this, they are considered as hermaphrodites. The life cycle of schistosomes includes two hosts: humans as definitive hosts, where the parasite undergoes sexual reproduction, and snails as intermediate hosts, where a series of asexual reproductive takes place. S. mansoni is transmitted through water, where freshwater snails of the genus Biomphalaria act as intermediate hosts. The larvae are able to live in water and infect the hosts by directly penetrating the skin. Prevention of infection is done by improved sanitation and killing the snails. Infection is treated with praziquantel. S. mansoni was first noted by Theodor Maximillian Bilharz in Egypt in 1851, while discovering S. haematobium. Sir Patrick Manson identified it as unique species in 1902.
After the eggs of the human-dwelling parasite are emitted in the faeces and into the water, the ripe miracidium hatches out of the egg. The hatching happens in response to temperature, light and dilution of faeces with water. The miracidium searches for a suitable freshwater snail belonging to the genus Biomphalaria. Miracidia directly penetrate the soft tissue of snail. Inside the snail, they lose their cilia and develop into mother sporocysts. The sporocysts rapidly multiply by asexual reproduction, each forming numerous daughter sporocysts. The daughter sporocysts move to the liver and gonads of the snail, where they undergo further growth. Within 2–4 weeks, they undergo metamorphosis and give rise to fork-tailed cercariae. Stimulated by light, hundreds of cercariae penetrate out of the snail into water.
The cercaria emerge from the snail during daylight and they propel themselves in water with the aid of their bifurcated tail, actively seeking out their final host. In water, they can live for up to 12 hours, and their maximum infectivity is between 1 and 9 hours after emergence. When they recognise human skin, they penetrate it within a very short time. This occurs in three stages, an initial attachment to the skin, followed by the creeping over the skin searching for a suitable penetration site, often a hair follicle, and finally penetration of the skin into the epidermis using cytolytic secretions from the cercarial post-acetabular, then pre-acetabular glands. On penetration, the head of the cercaria transforms into an endoparasitic larva, the schistosomule. Each schistosomule spends a few days in the skin and then enters the circulation starting at the dermal lymphatics and venules. Here, they feed on blood, regurgitating the haem as hemozoin. The schistosomule migrates to the lungs (5–7 days post-penetration) and then moves via circulation through the left side of the heart to the hepatoportal circulation (>15 days) where, if it meets a partner of the opposite sex, it develops into a sexually mature adult and the pair migrate to the mesenteric veins. Such pairings are monogamous.
Male schistosomes undergo normal maturation and morphological development in the presence or absence of a female, although behavioural, physiological and antigenic differences between males from single-sex, as opposed to bisex, infections have been reported. On the other hand, female schistosomes do not mature without a male. Female schistosomes from single-sex infections are underdeveloped and exhibit an immature reproductive system. Although the maturation of the female worm seems to be dependent on the presence of the mature male, the stimuli for female growth and for reproductive development seem to be independent from each other.
The adult female worm resides within the adult male worm’s gynaecophoric canal, which is a modification of the ventral surface of the male, forming a groove. The paired worms move against the flow of blood to their final niche in the mesenteric circulation, where they begin egg production (>32 days). The S. mansoni parasites are found predominantly in the small inferior mesenteric blood vessels surrounding the large intestine and caecal region of the host. Each female lays approximately 300 eggs a day (one egg every 4.8 minutes), which are deposited on the endothelial lining of the venous capillary walls. Most of the body mass of female schistosomes is devoted to the reproductive system. The female converts the equivalent of almost her own body dry weight into eggs each day. The eggs move into the lumen of the host’s intestines and are released into the environment with the faeces.
8.11 View Prepared Slides of Schistosoma
- Cercariae w.m. (Figure 8.10).
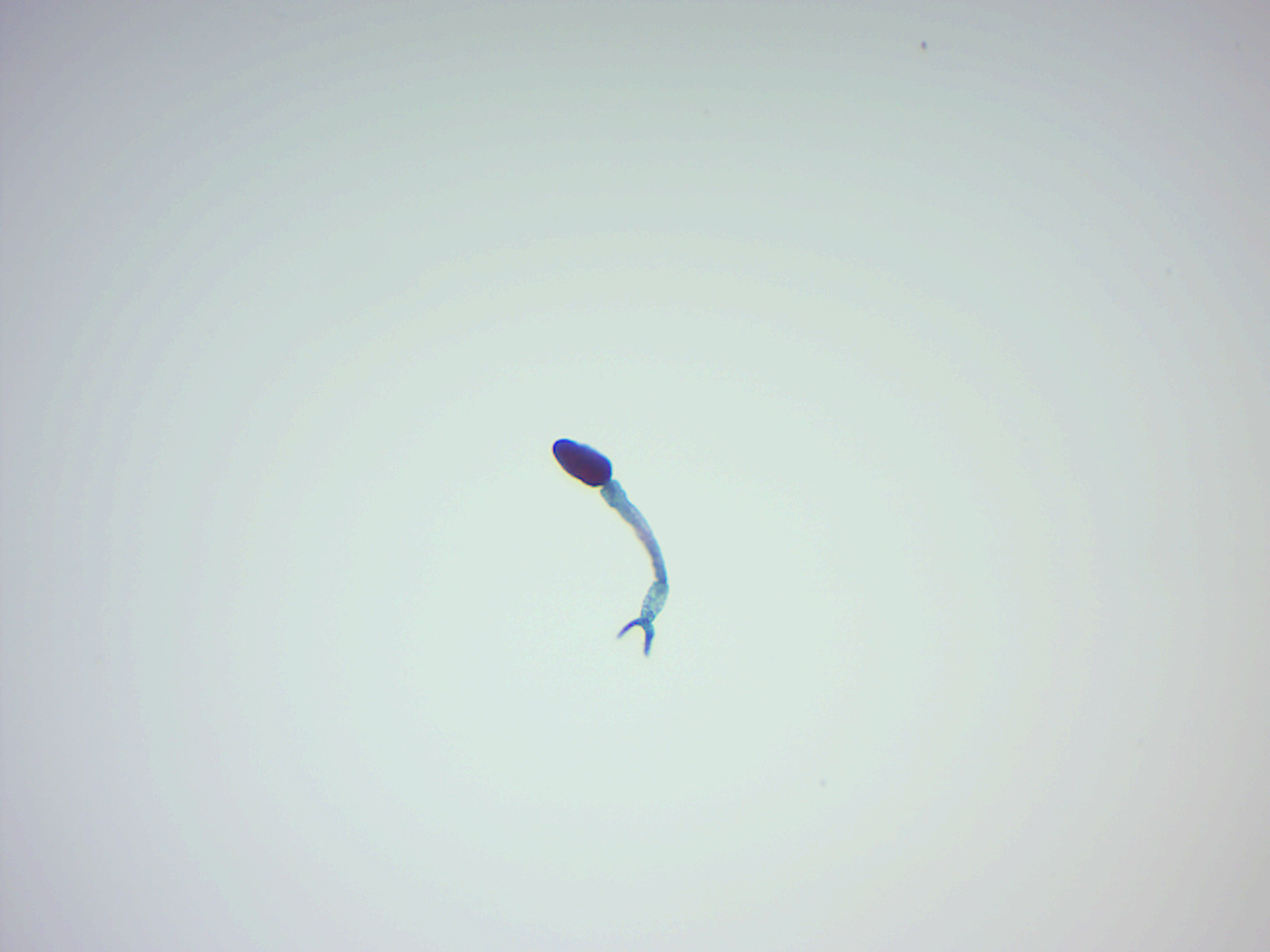
Figure 8.10: Schistosoma cercaria.
8.12 Monogenea
Monogeneans are a group of ectoparasite which are commonly found on the skin, gills, or fins of fish. They have a direct life cycle and do not require an intermediate host. Adults are hermaphrodites, meaning they have both male and female reproductive structures.
8.13 Molluska
Molluska are a large phylum of invertebrate animals whose members are known as mollusks. Around 85,000 extant species of mollusks are recognized. Mollusks are the largest marine phylum, comprising about 23% of all the named marine organisms. Numerous mollusks also live in freshwater and terrestrial habitats. They are highly diverse, not just in size and in anatomical structure, but also in behavior and in habitat. The phylum is typically divided into 9 or 10 taxonomic classes, of which two are entirely extinct. Cephalopod mollusks, such as squid, cuttlefish and octopus, are among the most neurologically advanced of all invertebrates—and either the giant squid or the colossal squid is the largest known invertebrate species. The gastropods (snails and slugs) are by far the most numerous mollusks in terms of classified species, and account for 80% of the total.
The three most universal features defining modern mollusks are a mantle with a significant cavity used for breathing and excretion, the presence of a radula (except for bivalves), and the structure of the nervous system. Other than these things, mollusks express great morphological diversity, so many textbooks base their descriptions on a “hypothetical ancestral mollusk” (see image below). This has a single, “limpet-like” shell on top, which is made of proteins and chitin reinforced with calcium carbonate, and is secreted by a mantle covering the whole upper surface. The underside of the animal consists of a single muscular “foot”. Although mollusks are coelomates, the coelom tends to be small. The main body cavity is a hemocoel through which blood circulates; their circulatory systems are mainly open. The “generalized” mollusk’s feeding system consists of a rasping “tongue”, the radula, and a complex digestive system in which exuded mucus and microscopic, muscle-powered “hairs” called cilia play various important roles. The generalized mollusk has two paired nerve cords, or three in bivalves. The brain, in species that have one, encircles the esophagus. Most mollusks have eyes, and all have sensors to detect chemicals, vibrations, and touch. The simplest type of molluskan reproductive system relies on external fertilization, but more complex variations occur. All produce eggs, from which may emerge trochophore larvae, more complex veliger larvae, or miniature adults.
Good evidence exists for the appearance of gastropods, cephalopods and bivalves in the Cambrian period 541 to 485.4 million years ago.
Mollusks have been and still are an important food source for anatomically modern humans. However there is a risk of food poisoning from toxins which can accumulate in certain mollusks under specific conditions, and because of this, many countries have regulations to reduce this risk. Mollusks have, for centuries, also been the source of important luxury goods, notably pearls, mother of pearl, Tyrian purple dye, and sea silk. Their shells have also been used as money in some preindustrial societies.
Mollusk species can also represent hazards or pests for human activities. Schistosomiasis (also known as bilharzia, bilharziosis or snail fever) is transmitted to humans via water snail hosts, and affects about 200 million people. Snails and slugs can also be serious agricultural pests, and accidental or deliberate introduction of some snail species into new environments has seriously damaged some ecosystems.
8.14 Bivalvia
Bivalves are a class of marine and freshwater mollusks that have laterally compressed bodies enclosed by a shell consisting of two hinged parts. Bivalves as a group have no head and they lack some usual molluskan organs like the radula and the odontophore (the cartilage which underlies and supports the radula and forms a ribbon of “teeth”). They include the clams, oysters, cockles, mussels, scallops, and numerous other families that live in saltwater, as well as a number of families that live in freshwater. The majority are filter feeders. The gills evolved into a specialized respiratory organ for feeding and breathing called ctenidium. Most bivalves bury themselves in sediment where they are relatively safe from predation. Others lie on the sea floor or attach themselves to rocks or other hard surfaces. Some bivalves, such as the scallops and file shells, can swim. The shipworms bore into wood, clay, or stone and live inside these substances.
The shell of a bivalve is composed of calcium carbonate, and consists of two, usually similar, parts called valves. These are joined together along one edge (the hinge line) by a flexible ligament that, usually in conjunction with interlocking “teeth” on each of the valves, forms the hinge. This arrangement allows the shell to be opened and closed without the two halves detaching. The shell is typically bilaterally symmetrical, with the hinge lying in the sagittal plane. Adult shell sizes of bivalves vary from fractions of a millimeter to over a meter in length, but the majority of species do not exceed 10 cm.
Bivalves appear in the fossil record first in the early Cambrian more than 500 million years ago. The total number of living species is about 9,200.
The sexes are usually separate in bivalves but some hermaphroditism is known. The gonads are located close to the intestines, and either open into the nephridia, or through a separate pore into the mantle cavity. The ripe gonads of male and females release sperm and eggs into the water column. Spawning may take place continually or be triggered by environmental factors such as day length, water temperature, or the presence of sperm in the water.
Fertilization is usually external. Typically, a short stage lasts a few hours or days before the eggs hatch into trochophore larvae. These later develop into veliger larvae which settle on the seabed and undergo metamorphosis into juveniles. In some species, such as those in the genus Lasaea, females draw water containing sperm in through their inhalant siphons and fertilization takes place inside the female. These species then brood the young inside their mantle cavity, eventually releasing them into the water column as veliger larvae or as crawl-away juveniles.
Most of the bivalve larvae that hatch from eggs in the water column feed on diatoms or other phytoplankton. In temperate regions, about 25% of species are lecithotrophic, depending on nutrients stored in the yolk of the egg where the main energy source is lipids. The larvae hatching out of these rely on the energy reserves and do not feed.
8.15 Clams
Clams have two shells of equal size connected by two adductor muscles and have a powerful burrowing foot. Some clams have life cycles of only one year, while at least one may be over 500 years old. All clams have two calcareous shells or valves joined near a hinge with a flexible ligament, and all are filter feeders. A clam’s shell consists of two (usually equal) valves, which are connected by a hinge joint and a ligament that can be external or internal. The ligament provides tension to bring the valves apart, while one or two adductor muscles can contract to close the valves. Clams also have kidneys, a heart, a mouth, a stomach, a nervous system and an anus. Many have a siphon.
Clams are protandrous hermaphrodites which means that they begin life as males but by the end of the first year approximately half of the population will change to females. Male clams release sperm into the water which stimulates females to expel eggs. A female may spawn several times each year, producing millions of eggs. Within 12 to 14 hours, the fertilized egg hatches into a microscopic creature called a trochophore larva (from the ancient Greek trókhos, meaning “wheel”, and phoréō, meaning “to bear, to carry” because the larva is bearing a wheel-shaped band of cilia). In less than a day, it transforms into a veliger larva (from Latin velum “sail, curtain, covering, veil” and -ger “bearing”), a free-swimming animal that has tiny wing-like lobes that propel it through the water. The foot, shell and body organs begin to form during the veliger stage, which lasts approximately 6 to 10 days. As the tiny shell develops, the veliger drops to the sea floor and sheds its lobes. When it touches bottom, it sends out thin filaments to hold it in place. As the clam matures, a muscular foot will replace these filaments, allowing the clam to bury itself in the sediments with only its siphons protruding.
8.16 Gastropoda
Gastropods (previously known as univalves) are a major part of the phylum Molluska, and are the most highly diversified class in the phylum, with 65,000 to 80,000 living snail and slug species.
In the older classification of the gastropods, there were four subclasses:
- Opisthobranchia (gills to the right and behind the heart).
- Gymnomorpha (no shell)
- Prosobranchia (gills in front of the heart).
- Pulmonata (with a lung instead of gills)
The class Gastropoda has an extraordinary diversification of habitats. Representatives live in gardens, woodland, deserts, and on mountains; in small ditches, great rivers and lakes; in estuaries, mudflats, the rocky intertidal, the sandy subtidal, in the abyssal depths of the oceans including the hydrothermal vents, and numerous other ecological niches, including parasitic ones.
Although the name “snail” can be, and often is, applied to all the members of this class, commonly this word means only those species with an external shell big enough that the soft parts can withdraw completely into it. Those gastropods without a shell, and those with only a very reduced or internal shell, are usually known as slugs; those with a shell into which they cannot withdraw are termed limpets.
The marine shelled species of gastropod include species such as abalone, conches, periwinkles, whelks, and numerous other sea snails that produce seashells that are coiled in the adult stage—though in some, the coiling may not be very visible, for example in cowries. In a number of families of species, such as all the various limpets, the shell is coiled only in the larval stage, and is a simple conical structure after that
Gastropods typically have a well-defined head with two or four sensory tentacles with eyes, and a ventral foot, which gives them their name (Greek gaster, stomach, and poda, feet). The upper pair of tentacles on the head of Helix pomatia have eye spots, but the main sensory organs of the snail are sensory receptors for olfaction, situated in the epithelium of the tentacles.
Sensory organs of gastropods include olfactory organs, eyes, statocysts and mechanoreceptors. Gastropods have no hearing. In terrestrial gastropods (land snails and slugs), the olfactory organs, located on the tips of the four tentacles, are the most important sensory organ. The majority of gastropods have simple visual organs, eye spots either at the tip or base of the tentacles. However, “eyes” in gastropods range from simple ocelli that only distinguish light and dark, to more complex pit eyes, and even to lens eyes. In land snails and slugs, vision is not the most important sense, because they are mainly nocturnal animals. The nervous system of gastropods includes the peripheral nervous system and the central nervous system. The central nervous system consists of ganglia connected by nerve cells.
The radula of a gastropod is usually adapted to the food that a species eats. The simplest gastropods are the limpets and abalones, herbivores that use their hard radula to rasp at seaweeds on rocks. Many marine gastropods are burrowers, and have a siphon that extends out from the mantle edge. Sometimes the shell has a siphonal canal to accommodate this structure. A siphon enables the animal to draw water into their mantle cavity and over the gill. They use the siphon primarily to “taste” the water to detect prey from a distance. Gastropods with siphons tend to be either predators or scavengers.
Almost all marine gastropods breathe with a gill, but many freshwater species, and the majority of terrestrial species, have a pallial lung.
Gastropods have open circulatory system and the transport fluid is hemolymph. Hemocyanin is present in the hemolymph as the respiratory pigment.
The primary organs of excretion in gastropods are nephridia, which produce either ammonia or uric acid as a waste product. The nephridium also plays an important role in maintaining water balance in freshwater and terrestrial species. Additional organs of excretion, at least in some species, include pericardial glands in the body cavity, and digestive glands opening into the stomach. In many marine gastropods other than the opisthobranchs, there are separate sexes; most land gastropods, however, are hermaphrodites.
8.17 View Prepared Slides of Molluska
- Snail Radula w.m. (Figure 8.11)
- Locate rows of chitinous teeth.
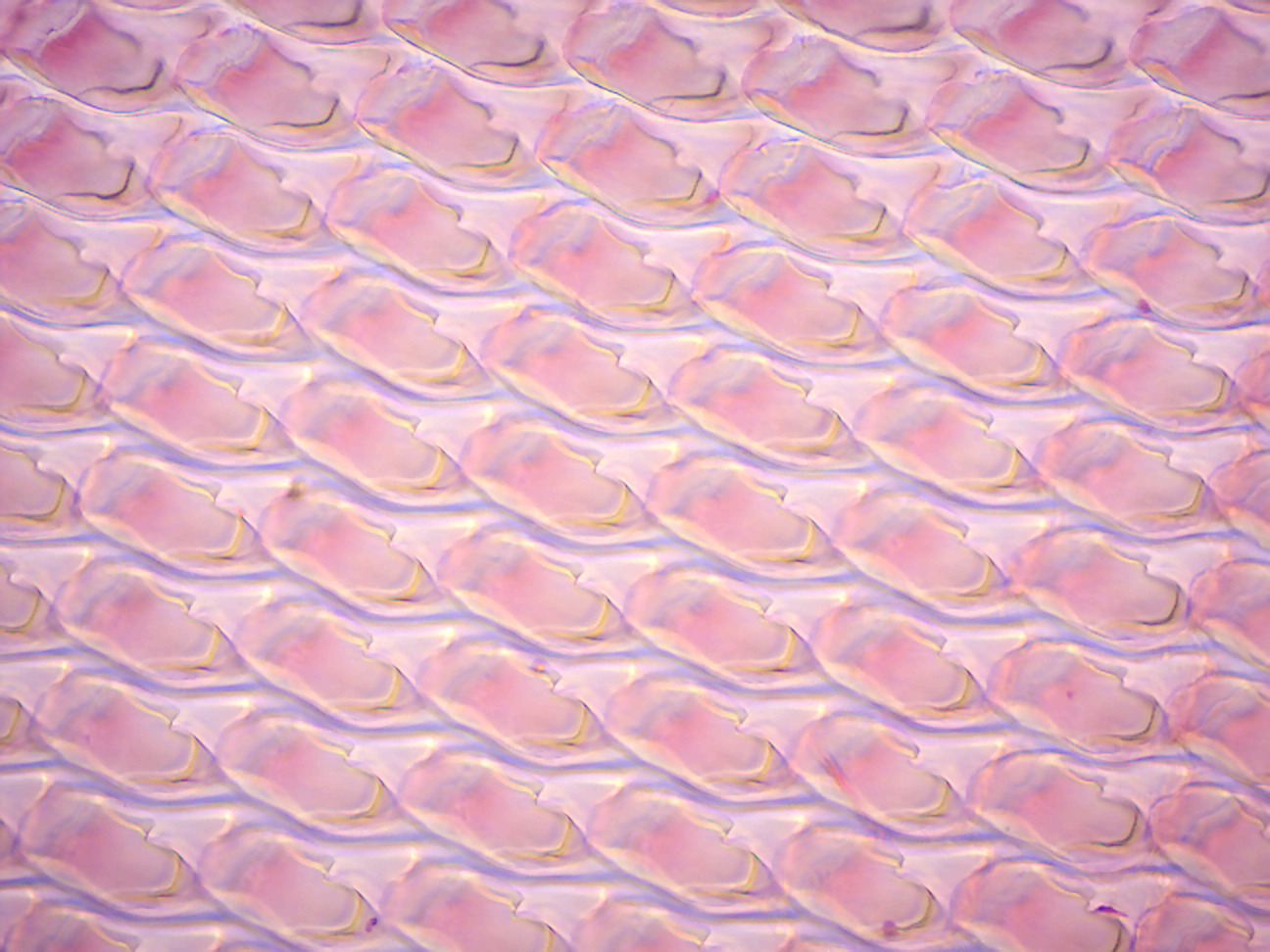
Figure 8.11: Snail radula.
8.18 Cephalopoda
Cephalopods are members of the molluskan class Cephalopoda (Greek plural kephalópoda; “head-feet”) such as a squid, octopus or nautilus. These exclusively marine animals are characterized by bilateral body symmetry, a prominent head, and a set of arms or tentacles modified from the primitive molluskan foot. About 800 living species of cephalopods have been identified. Two important extinct taxa are the Ammonoidea (ammonites) and Belemnoidea (belemnites).
Cephalopods are found in all the oceans of Earth. Cephalopods occupy most of the depth of the ocean, from the abyssal plain to the sea surface.
Cephalopods are widely regarded as the most intelligent of the invertebrates, and have well developed senses and large brains (larger than those of gastropods). The nervous system of cephalopods is the most complex of the invertebrates.
The brain is protected in a cartilaginous cranium. The giant nerve fibers of the cephalopod mantle have been widely used for many years as experimental material in neurophysiology; their large diameter (due to lack of myelination) makes them relatively easy to study compared with other animals. Many cephalopods are social creatures.
Cephalopods have advanced vision, can detect gravity with statocysts, and have a variety of chemical sense organs. Octopuses use their arms to explore their environment and can use them for depth perception.
Most cephalopods rely on vision to detect predators and prey, and to communicate with one another. Consequently, cephalopod vision is acute: training experiments have shown that the common octopus can distinguish the brightness, size, shape, and horizontal or vertical orientation of objects. The morphological construction gives cephalopod eyes the same performance as sharks’; however, their construction differs, as cephalopods lack a cornea, and have an everted retina. Cephalopods’ eyes are also sensitive to the plane of polarization of light. Surprisingly, given their ability to change color, all octopodes and most cephalopods are considered to be color blind. Coleoid cephalopods (octopus, squid, cuttlefish) have a single photoreceptor type and lack the ability to determine color by comparing detected photon intensity across multiple spectral channels. When camouflaging themselves, they use their chromatophores to change brightness and pattern according to the background they see, but their ability to match the specific color of a background may come from cells such as iridophores and leucophores that reflect light from the environment. They also produce visual pigments throughout their body, and may sense light levels directly from their body.
Cephalopods are the only mollusks with a closed circulatory system. Like most mollusks, cephalopods use hemocyanin, a copper-containing protein, rather than hemoglobin, to transport oxygen. As a result, their blood is colorless when deoxygenated and turns blue when exposed to air.
Cephalopods exchange gases with the seawater by forcing water through their gills, which are attached to the roof of the organism. Water enters the mantle cavity on the outside of the gills, and the entrance of the mantle cavity closes. When the mantle contracts, water is forced through the gills, which lie between the mantle cavity and the funnel. The water’s expulsion through the funnel can be used to power jet propulsion. The gills, which are much more efficient than those of other mollusks, are attached to the ventral surface of the mantle cavity.
All living cephalopods have a two-part beak; most have a radula, although it is reduced in most octopus. They feed by capturing prey with their tentacles, drawing it into their mouth and taking bites from it. They have a mixture of toxic digestive juices, some of which are manufactured by symbiotic algae, which they eject from their salivary glands onto their captured prey held in their mouth. These juices separate the flesh of their prey from the bone or shell. The salivary gland has a small tooth at its end which can be poked into an organism to digest it from within.
The digestive gland itself is rather short. It has four elements, with food passing through the crop, stomach and caecum before entering the intestine. Most digestion, as well as the absorption of nutrients, occurs in the digestive gland, sometimes called the liver. Nutrients and waste materials are exchanged between the gut and the digestive gland through a pair of connections linking the gland to the junction of the stomach and caecum. Cells in the digestive gland directly release pigmented excretory chemicals into the lumen of the gut, which are then bound with mucus passed through the anus as long dark strings, ejected with the aid of exhaled water from the funnel.
Most cephalopods possess a single pair of large nephridia. Filtered nitrogenous waste is produced in the pericardial cavity of the branchial hearts, each of which is connected to a nephridium by a narrow canal. The canal delivers the excreta to a bladder-like renal sac, and also resorbs excess water from the filtrate. Several outgrowths of the lateral vena cava project into the renal sac, continuously inflating and deflating as the branchial hearts beat. This action helps to pump the secreted waste into the sacs, to be released into the mantle cavity through a pore.
8.19 Annelida
The annelids (Annelida, from Latin anellus, “little ring”), also known as the ringed worms or segmented worms, are a large phylum, with over 17,000 extant species including ragworms, earthworms, and leeches. The species exist in and have adapted to various ecologies – some in marine environments as distinct as tidal zones and hydrothermal vents, others in fresh water, and yet others in moist terrestrial environments.
The annelids are bilaterally symmetrical, triploblastic, coelomate, invertebrate organisms. They also have parapodia for locomotion. Most textbooks still use the traditional division into polychaetes (almost all marine), oligochaetes (which include earthworms) and leech-like species. Cladistic research since 1997 has radically changed this scheme, viewing leeches as a sub-group of oligochaetes and oligochaetes as a sub-group of polychaetes. In addition, the Pogonophora, Echiura and Sipuncula, previously regarded as separate phyla, are now regarded as sub-groups of polychaetes. Annelids are considered members of the Lophotrochozoa, a “super-phylum” of protostomes that also includes mollusks’, brachiopods, flatworms and nemertea.
The basic annelid form consists of multiple segments. Each segment has the same sets of organs and, in most polychaetes, has a pair of parapodia that many species use for locomotion. Septa separate the segments of many species, but are poorly defined or absent in others, and Echiura and Sipuncula show no obvious signs of segmentation. In species with well-developed septa, the blood circulates entirely within blood vessels, and the vessels in segments near the front ends of these species are often built up with muscles that act as hearts. The septa of such species also enable them to change the shapes of individual segments, which facilitates movement by peristalsis (“ripples” that pass along the body) or by undulations that improve the effectiveness of the parapodia. In species with incomplete septa or none, the blood circulates through the main body cavity without any kind of pump, and there is a wide range of locomotory techniques – some burrowing species turn their pharynges inside out to drag themselves through the sediment.
Although many species can reproduce asexually and use similar mechanisms to regenerate after severe injuries, sexual reproduction is the normal method in species whose reproduction has been studied. The minority of living polychaetes whose reproduction and lifecycles are known produce trochophore larvae, that live as plankton and then sink and metamorphose into miniature adults. Oligochaetes are full hermaphrodites and produce a ring-like cocoon around their bodies, in which the eggs and hatchlings are nourished until they are ready to emerge.
Earthworms are Oligochaetes that support terrestrial food chains both as prey and in some regions, are important in aeration and enriching of soil. The burrowing of marine polychaetes, which may constitute up to a third of all species in near-shore environments, encourages the development of ecosystems by enabling water and oxygen to penetrate the sea floor. In addition to improving soil fertility, annelids serve humans as food and as bait. Scientists observe annelids to monitor the quality of marine and fresh water. Although blood-letting is no longer in favor with doctors, some leech species are regarded as endangered species because they have been over-harvested for this purpose in the last few centuries.
Since annelids are soft-bodied, their fossils are rare – mostly jaws and the mineralized tubes that some of the species secreted. Although some late Ediacaran fossils may represent annelids, the oldest known fossil that is identified with confidence comes from about 518 million years ago in the early Cambrian period. Fossils of most modern mobile polychaete groups appeared by the end of the Carboniferous, about 299 million years ago. Palaeontologists disagree about whether some body fossils from the mid Ordovician, about 472 to 461 million years ago, are the remains of oligochaetes, and the earliest indisputable fossils of the group appear in the Tertiary period, which began 65 million years ago.
8.20 Earthworm
An earthworm is a tube-shaped, segmented worm found in the phylum Annelida. Earthworms are commonly found living in soil, feeding on live and dead organic matter. An earthworm’s digestive system runs through the length of its body. It conducts respiration through its skin. It has a double transport system composed of coelomic fluid that moves within the fluid-filled coelom and a simple, closed blood circulatory system. It has a central and a peripheral nervous system. The central nervous system consists of two ganglia above the mouth, one on either side, connected to a nerve cord running back along its length to motor neurons and sensory cells in each segment. Large numbers of chemoreceptors are concentrated near its mouth. Circumferential and longitudinal muscles on the periphery of each segment enable the worm to move. Similar sets of muscles line the gut, and their actions move the digesting food toward the worm’s anus.
Earthworms are hermaphrodites–each individual carries both male and female sex organs. They lack either an internal skeleton or exoskeleton, but maintain their structure with fluid-filled coelom chambers that function as a hydrostatic skeleton.
Depending on the species, an adult earthworm can be from 10 mm long and 1 mm wide to 3 m long and over 25 mm wide, but the typical Lumbricus terrestris grows to about 360 mm long. Probably the longest worm on confirmed records is Amynthas mekongianus that extends up to 3 m in the mud along the banks of the Mekong River in Southeast Asia.
From front to back, the basic shape of the earthworm is a cylindrical tube, divided into a series of segments (called metamerisms) that compartmentalize the body. Furrows are generally externally visible on the body demarking the segments; dorsal pores and nephridiopores exude a fluid that moistens and protects the worm’s surface, allowing it to breathe. Except for the mouth and anal segments, each segment carries bristle-like hairs called lateral setae used to anchor parts of the body during movement; species may have four pairs of setae on each segment or more than eight sometimes forming a complete circle of setae per segment.
Generally, within a species, the number of segments found is consistent across specimens, and individuals are born with the number of segments they will have throughout their lives. The first body segment (segment number 1) features both the earthworm’s mouth and, overhanging the mouth, a fleshy lobe called the prostomium, which seals the entrance when the worm is at rest, but is also used to feel and chemically sense the worm’s surroundings. Some species of earthworm can even use the prehensile prostomium to grab and drag items such as grasses and leaves into their burrow.
An adult earthworm develops a belt-like glandular swelling, called the clitellum, which covers several segments toward the front part of the animal. This is part of the reproductive system and produces egg capsules. The posterior is most commonly cylindrical like the rest of the body, but depending on the species, may also be quadrangular, octagonal, trapezoidal, or flattened. The last segment is called the periproct; the earthworm’s anus, a short vertical slit, is found on this segment.
The exterior of an individual segment is a thin cuticle over skin, commonly pigmented red to brown, which has specialized cells that secrete mucus over the cuticle to keep the body moist and ease movement through soil. Under the skin is a layer of nerve tissue, and two layers of muscles—a thin outer layer of circular muscle, and a much thicker inner layer of longitudinal muscle. Interior to the muscle layer is a fluid-filled chamber called a coelom that by its pressurization provides structure to the worm’s boneless body. The segments are separated from each other by septa (the plural of “septum”) which are perforated transverse walls, allowing the coelomic fluid to pass between segments. A pair of structures called nephrostomes are located at the back of each septum; a nephric tubule leads from each nephrostome through the septum and into the following segment. This tubule then leads to the main body fluid filtering organ, the nephridium or metanephridium, which removes metabolic waste from the coelomic fluid and expels it through pores called nephridiopores on the worm’s sides; usually two nephridia (sometimes more) are found in most segments. At the center of a worm is the digestive tract, which runs straight through from mouth to anus without coiling, and is flanked above and below by blood vessels (the dorsal blood vessel and the ventral blood vessel as well as a subneural blood vessel) and the ventral nerve cord, and is surrounded in each segment by a pair of pallial blood vessels that connect the dorsal to the subneural blood vessels.
The earthworm’s nervous system has three parts: the central nervous system (CNS), peripheral nervous system and the sympathetic nervous system.
The gut of the earthworm is a straight tube which extends from the worm’s mouth to its anus. It is differentiated into a buccal cavity (generally running through the first one or two segments of the earthworm), pharynx (running generally about four segments in length), esophagus, crop, gizzard (usually) and intestine. Food enters in the mouth. The pharynx acts as a suction pump; its muscular walls draw in food. In the pharynx, the pharyngeal glands secrete mucus. Food moves into the esophagus. From there the food passes into the crop and gizzard. In the gizzard, strong muscular contractions grind the food with the help of mineral particles ingested along with the food. Once through the gizzard, food continues through the intestine for digestion. Instead of being coiled like a mammalian intestine, an earthworm’s intestine increases surface area to increase nutrient absorption by having many folds running along its length. The intestine has its own pair of muscle layers like the body, but in reverse order—an inner circular layer within an outer longitudinal layer.
The earthworm has a dual circulatory system in which both the coelomic fluid and a closed circulatory system carry the food, waste, and respiratory gases. The closed circulatory system has five main blood vessels: the dorsal (top) vessel, which runs above the digestive tract; the ventral (bottom) vessel, which runs below the digestive tract; the subneural vessel, which runs below the ventral nerve cord; and two lateroneural vessels on either side of the nerve cord. The dorsal vessel moves the blood forward, while the other four longitudinal vessels carry the blood rearward. In segments seven through eleven, a pair of aortic arches rings the coelom and acts as hearts, pumping the blood to the ventral vessel that acts as the aorta. The blood consists of ameboid cells and hemoglobin dissolved in the plasma. The second circulatory system derives from the cells of the digestive system that line the coelom.
The excretory system contains a pair of nephridia in every segment, except for the first three and the last ones. The waste in the coelom fluid from a forward segment is drawn in by the beating of cilia of the nephrostome. From there it is carried through the septum (wall) via a tube which forms a series of loops entwined by blood capillaries that also transfer waste into the tubule of the nephrostome. The excretory wastes are then finally discharged through a pore on the worm’s side.
Earthworms have no special respiratory organs. Gases are exchanged through the moist skin and capillaries, where the oxygen is picked up by the hemoglobin dissolved in the blood plasma and carbon dioxide is released. Water, as well as salts, can also be moved through the skin by active transport.
Mating occurs on the surface, most often at night. Earthworms are hermaphrodites; that is, they have both male and female sexual organs. The sexual organs are located in segments 9 to 15. Earthworms have one or two pairs of testes contained within sacs. The two or four pairs of seminal vesicles produce, store and release the sperm via the male pores. Ovaries and oviducts in segment 13 release eggs via female pores on segment 14, while sperm is expelled from segment 15. One or more pairs of spermathecae are present in segments 9 and 10 (depending on the species) which are internal sacs that receive and store sperm from the other worm during copulation. As a result, segment 15 of one worm exudes sperm into segments 9 and 10 with its storage vesicles of its mate. Some species use external spermatophores for sperm transfer. Copulation and reproduction are separate processes in earthworms. The mating pair overlap front ends ventrally and each exchanges sperm with the other. The clitellum becomes very reddish to pinkish in color. Sometime after copulation, long after the worms have separated, the clitellum (behind the spermathecae) secretes material which forms a ring around the worm. The worm then backs out of the ring, and as it does so, it injects its own eggs and the other worm’s sperm into it. As the worm slips out of the ring, the ends of the cocoon seal to form a vaguely lemon-shaped incubator (cocoon) in which the embryonic worms develop. They emerge as small, but fully formed earthworms, but lack their sex structures, which develop in about 60 to 90 days. They attain full size in about one year. Scientists predict that the average lifespan under field conditions is four to eight years, while most garden varieties live only one to two years.
8.21 View Living Organisms
- Turbellaria
- Rotifers
Please watch the following video of a clam dissection:
8.22 Clam Dissection
- Get a dissecting pan, a scalpel, forceps, and a pointer.
- Get a clam.
- Identify the anterior and posterior ends of the clam as well as the dorsal, ventral, and left and right lateral surfaces (Figure 8.12).
- Place a clam in a dissecting tray with the left side up facing you.
- Locate the umbo, the bump at the anterior end of the valve. This is the oldest part of the clam shell. Find the hinge ligament which hinges the valves together and observe the growth rings.
- Locate the adductor muscles. With your blade pointing toward the dorsal edge, slide your scalpel between the two valves & and cut through the anterior adductor muscle, cutting as close to the shell as possible.
- Repeat step 6 in cutting the posterior adductor muscle.
- Bend the left valve back so it lies flat on the tray. Once both adductor muscles have been cut away from the left shell, the shell can be opened easily.
- Locate the muscle “scars” on the inner surface of the left valve. The adductor muscles were attached here to hold the clam closed.
- Identify the mantle, the tissue that lines both valves & covers the soft body of the clam. Find the mantle cavity, the space inside the mantle (see Figure 8.13).
- Locate two openings on the posterior end of the clam. The more ventral opening is the incurrent siphon that carries water into the clam and the more dorsal opening is the excurrent siphon where wastes and water leave. The large muscle attached to the siphons is called the siphon retractor muscle. There would be another one on the right side. These muscles pull the siphon in. Most clams can retract the siphons completely into the shell.
- Lift the mantle up. Notice how thin it is. Remove the mantle to expose the gills and the foot. Notice the structure of the gills. Bivalves are filter feeders. The gills are used to strain plankton out of the water, as well as to remove oxygen from the water. There are another pair of gills on the right side of the clam.
- Observe the muscular foot of the clam, which is ventral to the gills. Note the hatchet shape of the foot used to burrow into mud or sand.
- Locate the labial palps, flaplike structures that surround and guide food into the clam’s mouth. The palps are anterior to the gills & ventral to the anterior adductor muscle. Between the palps, find the mouth (see Figure 8.14).
- Carefully make an incision in the middle of the visceral mass to view the internal organs.
- Locate the spongy, yellowish reproductive organs.
- Ventral to the umbo, find the digestive gland, a greenish structure that surrounds the stomach.
- Locate the long, coiled intestine extending from the stomach.
- Follow the intestine through the clam. Find the area near the dorsal surface that the intestine passes through called the pericardial area. Find the clam’s heart in this area (see Figure 8.15).
- Continue following the intestine toward the posterior end of the clam. The heart is contained in a thin-walled sac called the pericardium near the top of the clam above the visceral mass. The tube-like structure that runs through the pericardium is the intestine.

Figure 8.12: External features of the clam. Lateral view of the left side of a clam shell. Note the umbo (top left) and growth rings.
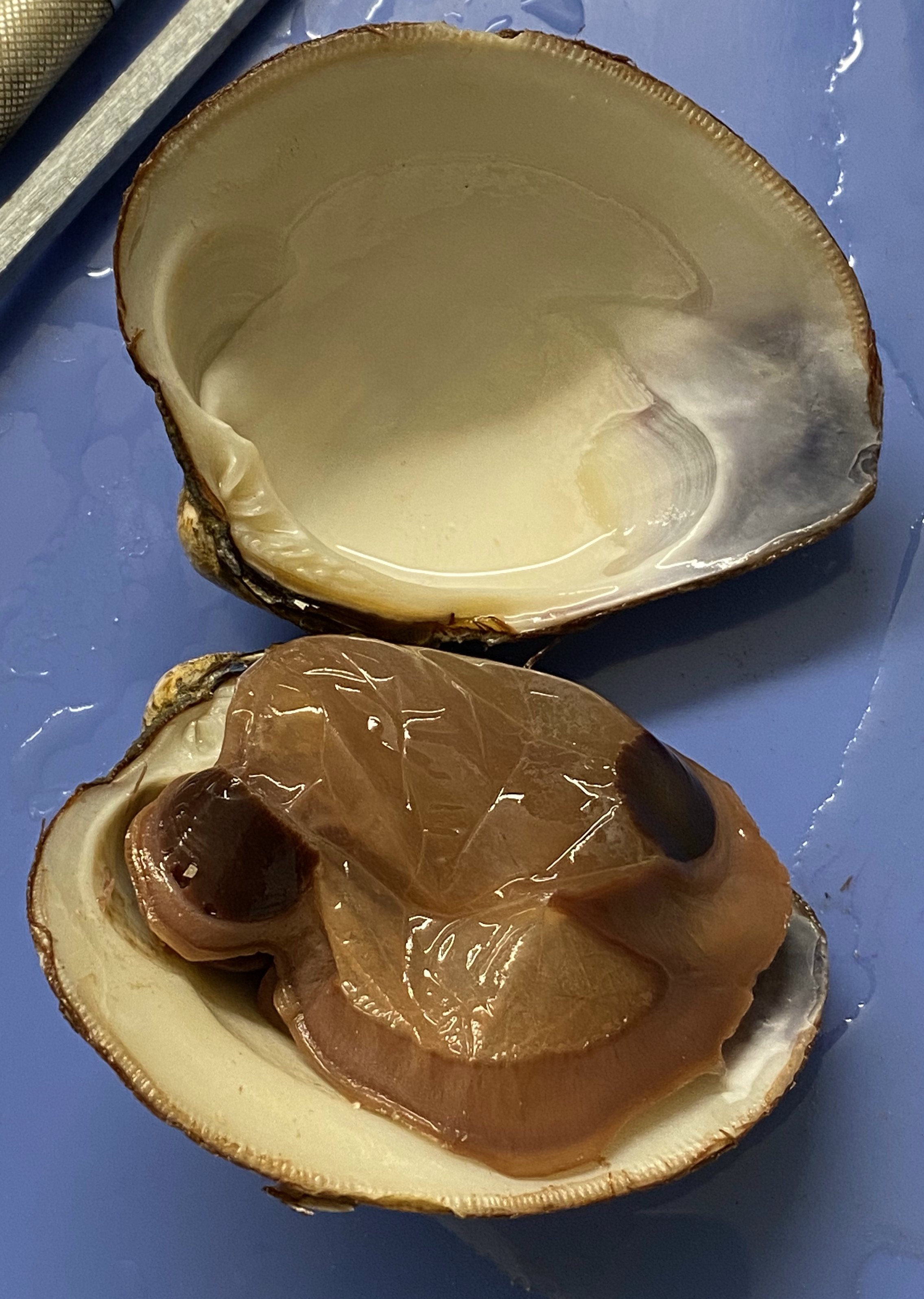
Figure 8.13: View of the mantle covering the left side of the clam.
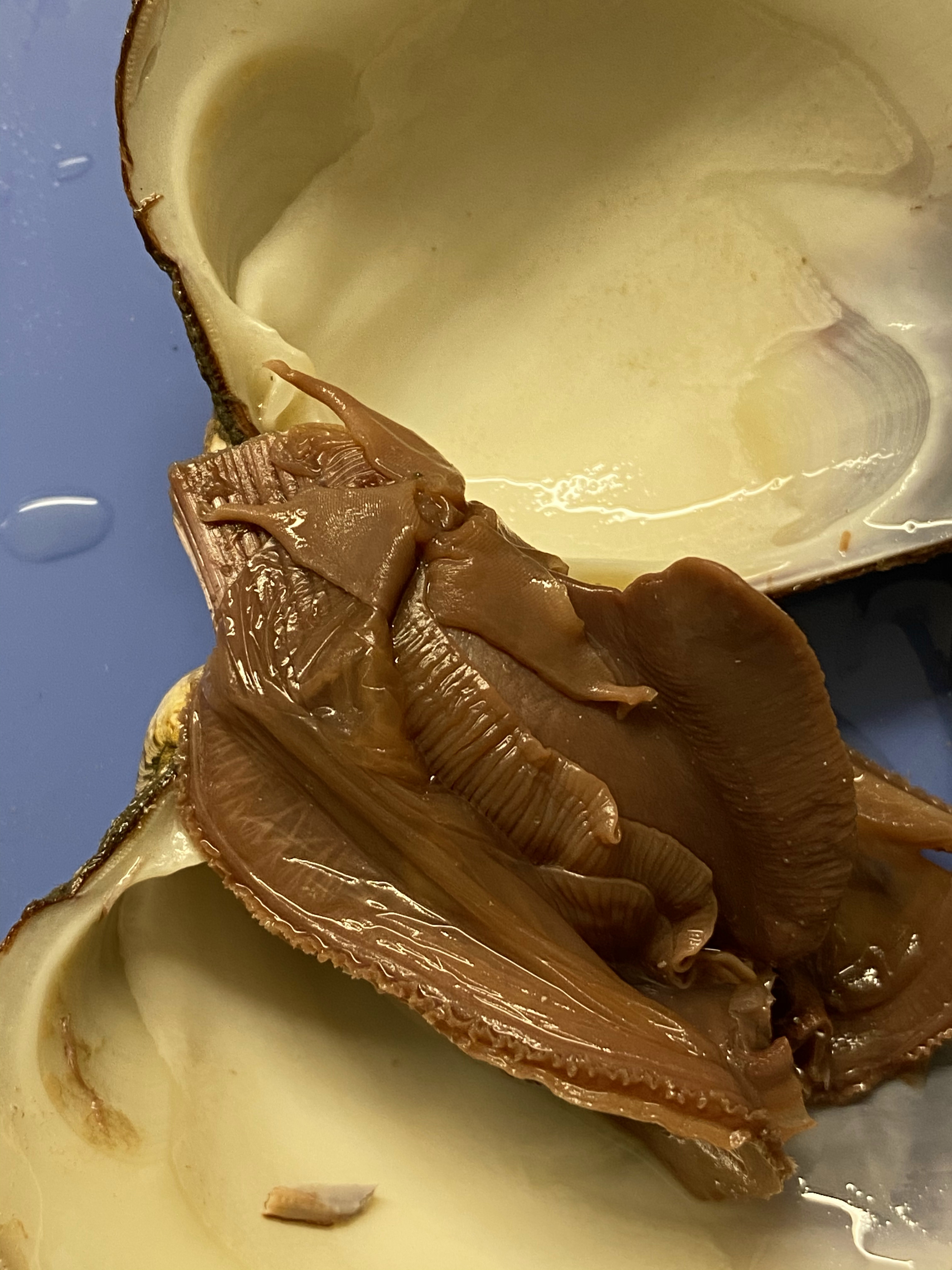
Figure 8.14: View of the left side of the clam after removal of the mantle. Note the labial palps, foot and gills.
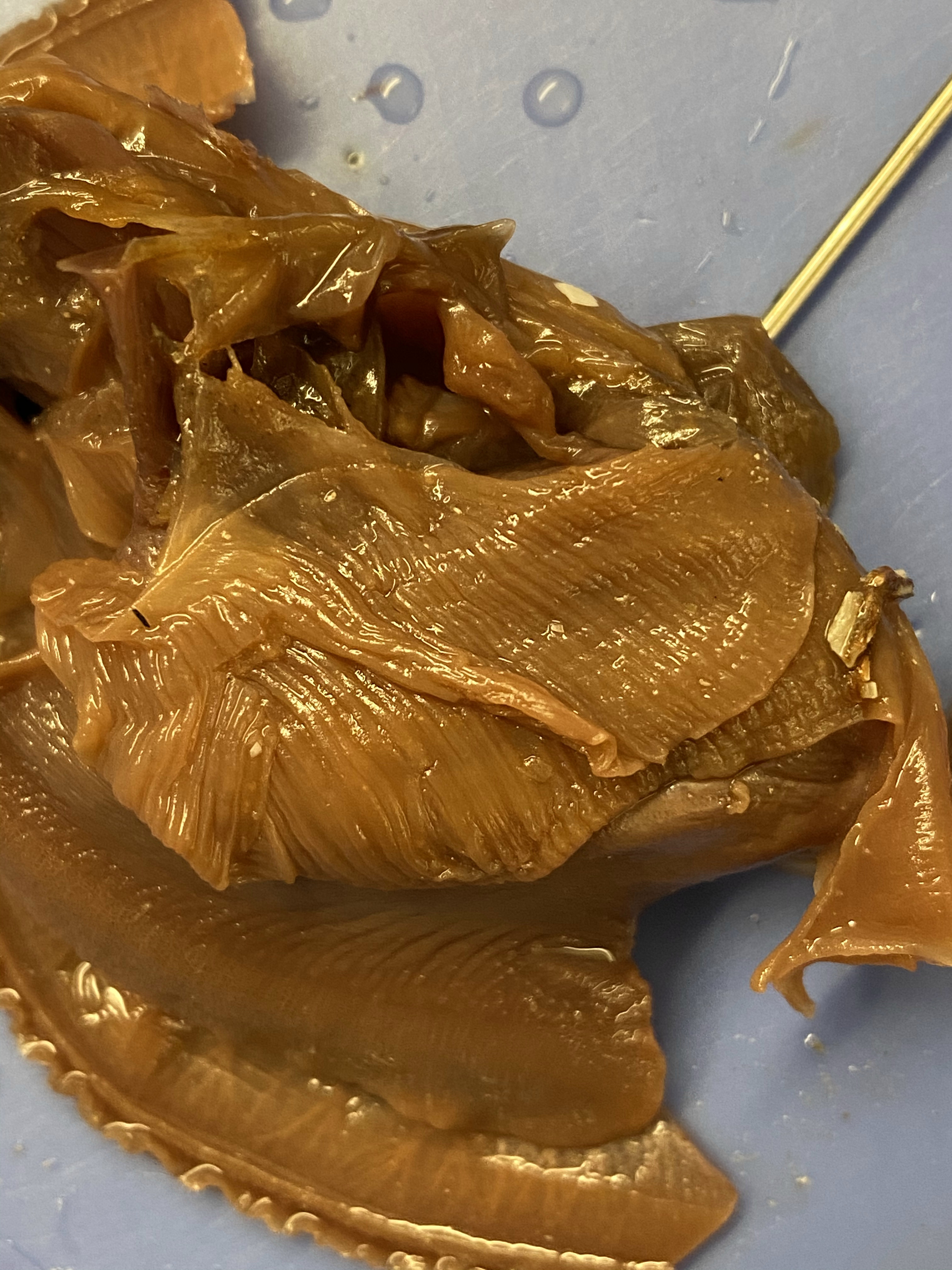
Figure 8.15: View of the coelomic cavity with the heart exposed.
8.23 Cleaning up
- Dispose of the remains of the clam in the red biohazard bins.
- Clean the dissection tray and instruments.
8.24 Earthworm Dissection
- Get a dissecting pan, a scalpel, forceps, and a pointer.
- Get an earhtworm.
- Position your preserved earthworm dorsal side up and pin it down through the first segment and then again further back behind the clitellum (Figure 8.16).
- Cut a slit in the dorsal surface near the posterior pin and extend the cut forward to the first segment. Be careful not to cut too deep.
- Starting at the first segment, cut the septa (thin membranes) that internally divide the segments, so the skin can be laid flat. Use additional pins to hold the integument open and expose the organs.
- Continue to lay the skin back until you have uncovered a centimeter or so of the intestine.
- Locate and identify the five pairs of aortic arches, or hearts. Then find the dorsal blood vessel. Look for smaller blood vessels that branch from the dorsal blood vessel.
- Starting at the anterior end, locate the muscular pharynx (food ingestion). This is followed by a tube-like esophagus which terminates in a crop (the wider organ) which serves as a storage stomach. Posterior to the crop is the gizzard. Gently press on the crop and gizzard to test their firmness. While the crop is soft and thin, the gizzard is muscular (soil is ground up and churned within the gizzard). The gizzard is followed by a long intestine in which both digestion and absorption occur. Undigested material is voided through the anus.
- To find organs of the nervous system, push aside the digestive and circulatory system organs. Locate the ventral nerve cord. Trace the nerve cord forward to the nerve collar, which circles the pharynx. Find one pair of ganglia under the pharynx and another pair of ganglia above the pharynx. The ganglia above the pharynx serve as the brain of the earthworm.
- Locate the nephridia. There are two in every segment.
- Locate and identify a pair of ovaries in segment 13.
- Look for two pairs of tiny testes in segments 10 and 11. To find these organs, you will again have to push aside some parts already dissected.
8.25 Cleaning up
- Dispose of the remains of the earthworm in the red biohazard bins.
- Clean the dissection tray and instruments and return them to the place where you picked them up.
- Clean table tops with red bottled sanitizer
- Wash hands before leaving class.
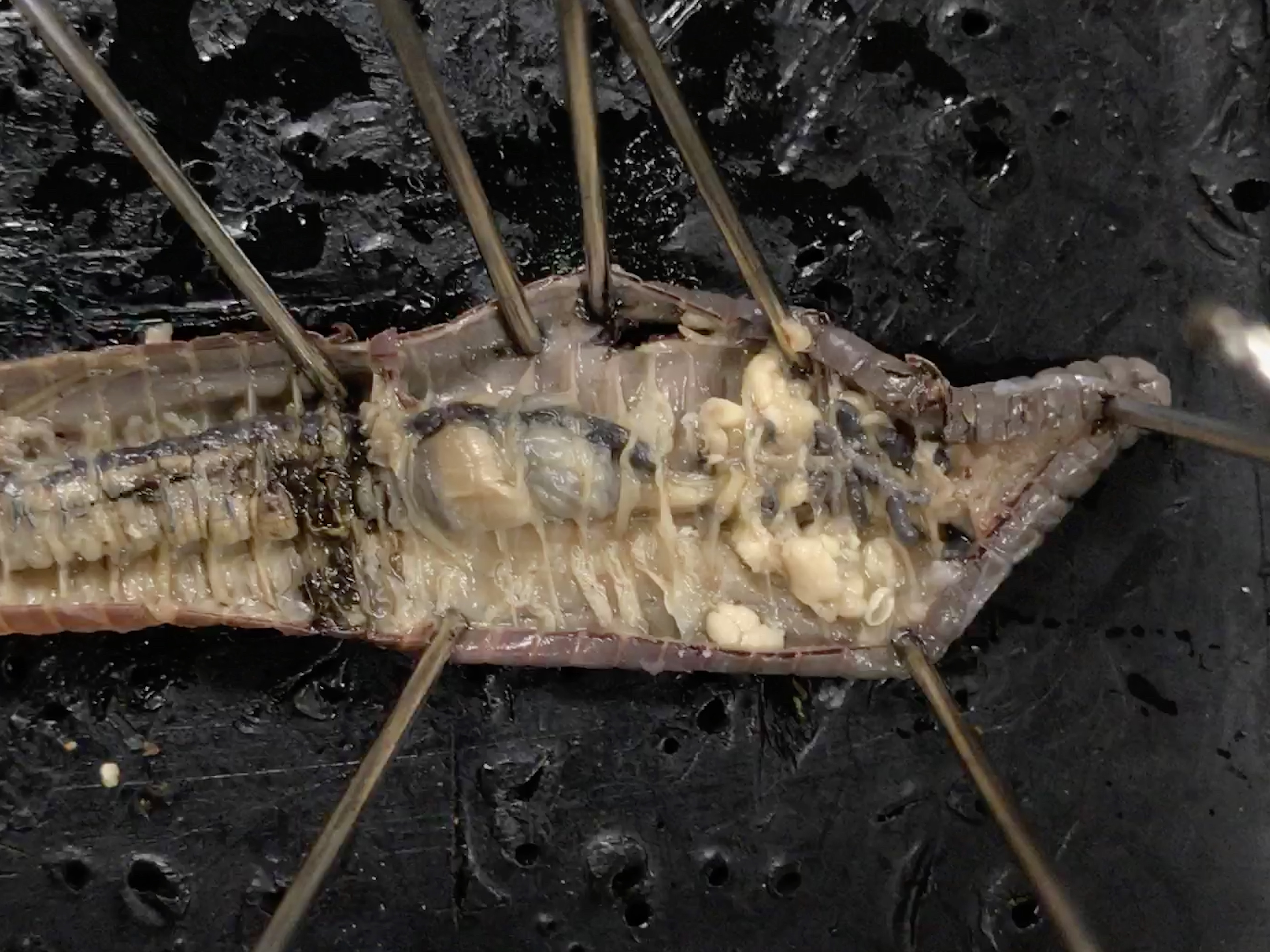
Figure 8.16: Earthworm dissection
8.26 Review Questions
- What are rotifers?
- What are flat worms?
- What are mollusks?
- What are annelids?
- What is segmentation?
- What is a radula?
- What is a closed circulatory system?
- What is an open circulatory system?