20 Circulatory Systems
The circulatory system, also called the cardiovascular system or the vascular system, is an organ system that permits blood to circulate and transport nutrients (such as amino acids and electrolytes), oxygen, carbon dioxide, hormones, and blood cells to and from the cells in the body to provide nourishment and help in fighting diseases, stabilize temperature and pH, and maintain homeostasis. The cardiovascular (from Latin words meaning “heart” and “vessel”) system comprises the blood, heart, and blood vessels.
While humans, as well as other vertebrates, have a closed cardiovascular system (meaning that the blood never leaves the network of arteries, veins and capillaries), some invertebrate groups have an open cardiovascular system. The more primitive, diploblastic animal phyla lack circulatory systems.
20.1 Closed Circulatory System
The circulatory systems of all vertebrates, as well as of annelids (for example, earthworms) and cephalopods (squids, octopuses and relatives) are closed, just as in humans. Still, the systems of fish, amphibians, reptiles, and birds show various stages of the evolution of the circulatory system.
Fish have the simplest circulatory system. It has only one circuit, with the blood being pumped through the capillaries of the gills and on to the capillaries of the body tissues. This is known as single cycle circulation. The heart of fish is, therefore, only a single pump consisting of one atrium to receive blood and one ventricle to pump it, in contrast to three chambers (two atria, one ventricle) of amphibian and most reptile hearts and four chambers (two atria, two ventricles) of mammal and bird hearts. However, the fish heart has entry and exit compartments that may be called chambers, so it is also sometimes described as three-chambered or four-chambered, depending on what is counted as a chamber. The atrium and ventricle are sometimes considered “true chambers”, while the others are considered “accessory chambers”.
The four compartments are arranged sequentially:
- Sinus venosus, a thin-walled sac or reservoir with some cardiac muscle that collects deoxygenated blood through the incoming hepatic and cardinal veins.
- Atrium, a thicker-walled, muscular chamber that sends blood to the ventricle.
- Ventricle, a thick-walled, muscular chamber that pumps the blood to the fourth part, the outflow tract. The shape of the ventricle varies considerably, usually tubular in fish with elongated bodies, pyramidal with a triangular base in others, or sometimes sac-like in some marine fish.
- The outflow tract (OFT) to the ventral aorta, consisting of the tubular conus arteriosus, bulbus arteriosus, or both. The conus arteriosus, typically found in more primitive species of fish, contracts to assist blood flow to the aorta, while the bulbus anteriosus does not.
Ostial valves, consisting of flap-like connective tissues, prevent blood from flowing backward through the compartments. The ostial valve between the sinus venosus and atrium is called the sino-atrial valve, which closes during ventricular contraction. Between the atrium and ventricle is an ostial valve called the atrio-ventricular valve, and between the bulbus arteriosus and ventricle is an ostial valve called the bulbo-ventricular valve. The conus arteriosus has a variable number of semilunar valves.
The ventral aorta delivers blood to the gills where it is oxygenated and flows, through the dorsal aorta, into the rest of the body. (In tetrapods, the ventral aorta has divided in two; one half forms the ascending aorta, while the other forms the pulmonary artery).
In amphibians and most reptiles, a double circulatory system is used, but the heart is not always completely separated into two pumps. Amphibians have a three-chambered heart.
In reptiles, the ventricular septum of the heart is incomplete and the pulmonary artery is equipped with a sphincter muscle. This allows a second possible route of blood flow. Instead of blood flowing through the pulmonary artery to the lungs, the sphincter may be contracted to divert this blood flow through the incomplete ventricular septum into the left ventricle and out through the aorta. This means the blood flows from the capillaries to the heart and back to the capillaries instead of to the lungs. This process is useful to ectothermic (cold-blooded) animals in the regulation of their body temperature.
Birds, mammals, and crocodilians show complete separation of the heart into two pumps, for a total of four heart chambers; it is thought that the four-chambered heart of birds and crocodilians evolved independently from that of mammals.
20.2 Open Circulatory Systems
In arthropods, the open circulatory system is a system in which a fluid in a cavity called the hemocoel bathes the organs directly with oxygen and nutrients and there is no distinction between blood and interstitial fluid; this combined fluid is called hemolymph or haemolymph. Muscular movements by the animal during locomotion can facilitate hemolymph movement, but diverting flow from one area to another is limited. When the heart relaxes, blood is drawn back toward the heart through open-ended pores (ostia).
Hemolymph fills all of the interior hemocoel of the body and surrounds all cells. Hemolymph is composed of water, inorganic salts (mostly sodium, chloride, potassium, magnesium, and calcium), and organic compounds (mostly carbohydrates, proteins, and lipids). The primary oxygen transporter molecule is hemocyanin.
There are free-floating cells, the hemocytes, within the hemolymph. They play a role in the arthropod immune system.
20.3 No Circulatory System
Circulatory systems are absent in some animals, including flatworms. Their body cavity has no lining or enclosed fluid. Instead a muscular pharynx leads to an extensively branched digestive system that facilitates direct diffusion of nutrients to all cells. The flatworm’s dorso-ventrally flattened body shape also restricts the distance of any cell from the digestive system or the exterior of the organism. Oxygen can diffuse from the surrounding water into the cells, and carbon dioxide can diffuse out. Consequently, every cell is able to obtain nutrients, water and oxygen without the need of a transport system.
Some animals, such as jellyfish, have more extensive branching from their gastrovascular cavity (which functions as both a place of digestion and a form of circulation), this branching allows for bodily fluids to reach the outer layers, since the digestion begins in the inner layers.
20.4 The Human Cardiovascular System
The essential components of the human cardiovascular system are the heart, blood and blood vessels. Blood is a fluid consisting of plasma, red blood cells, white blood cells, and platelets that is circulated by the heart through the vertebrate vascular system, carrying oxygen and nutrients to and waste materials away from all body tissues. The circulatory system of the blood is seen as having two components, a systemic circulation and a pulmonary circulation.
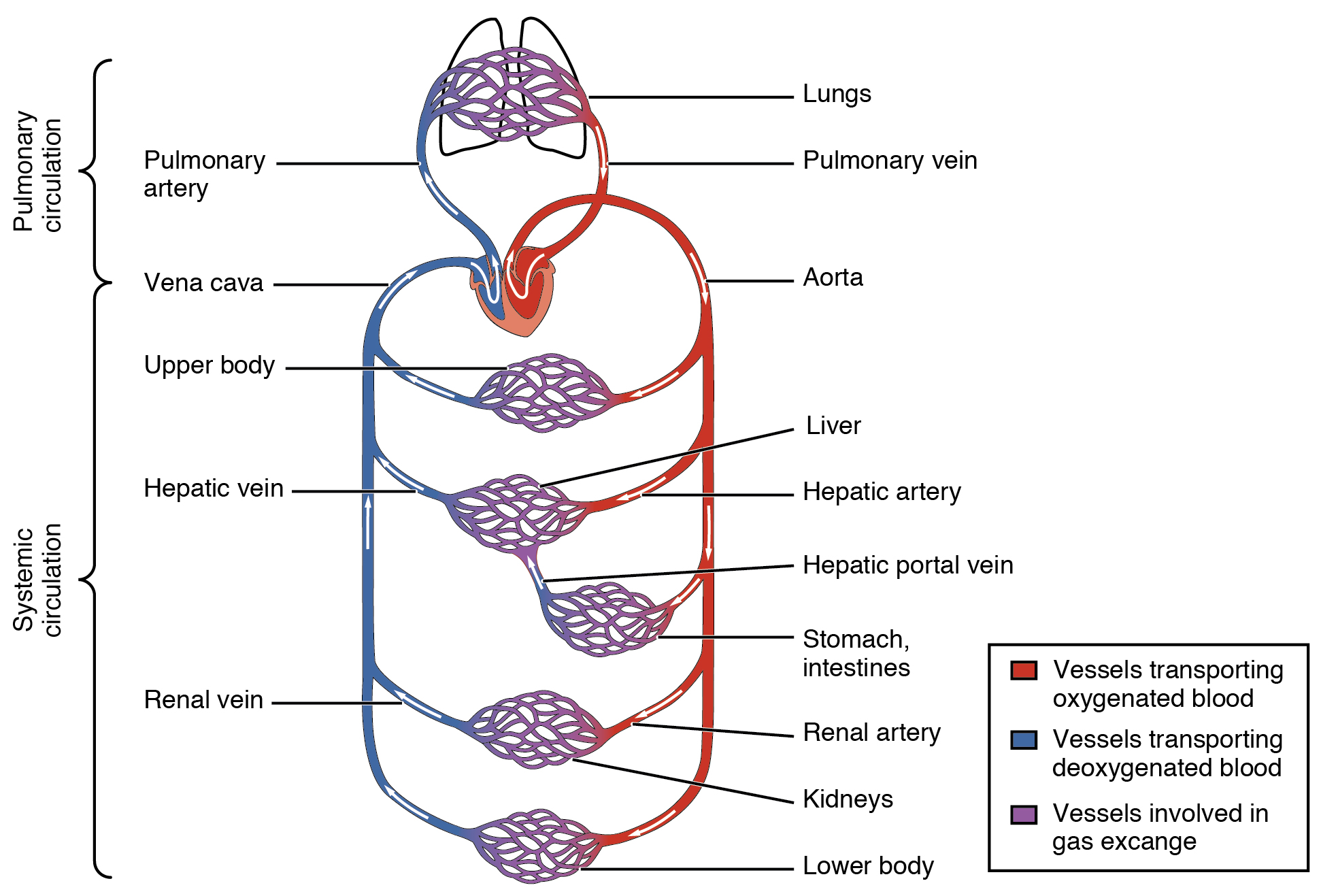
Figure 20.1: The systemic circulation and capillary networks shown and also as separate from the pulmonary circulation.
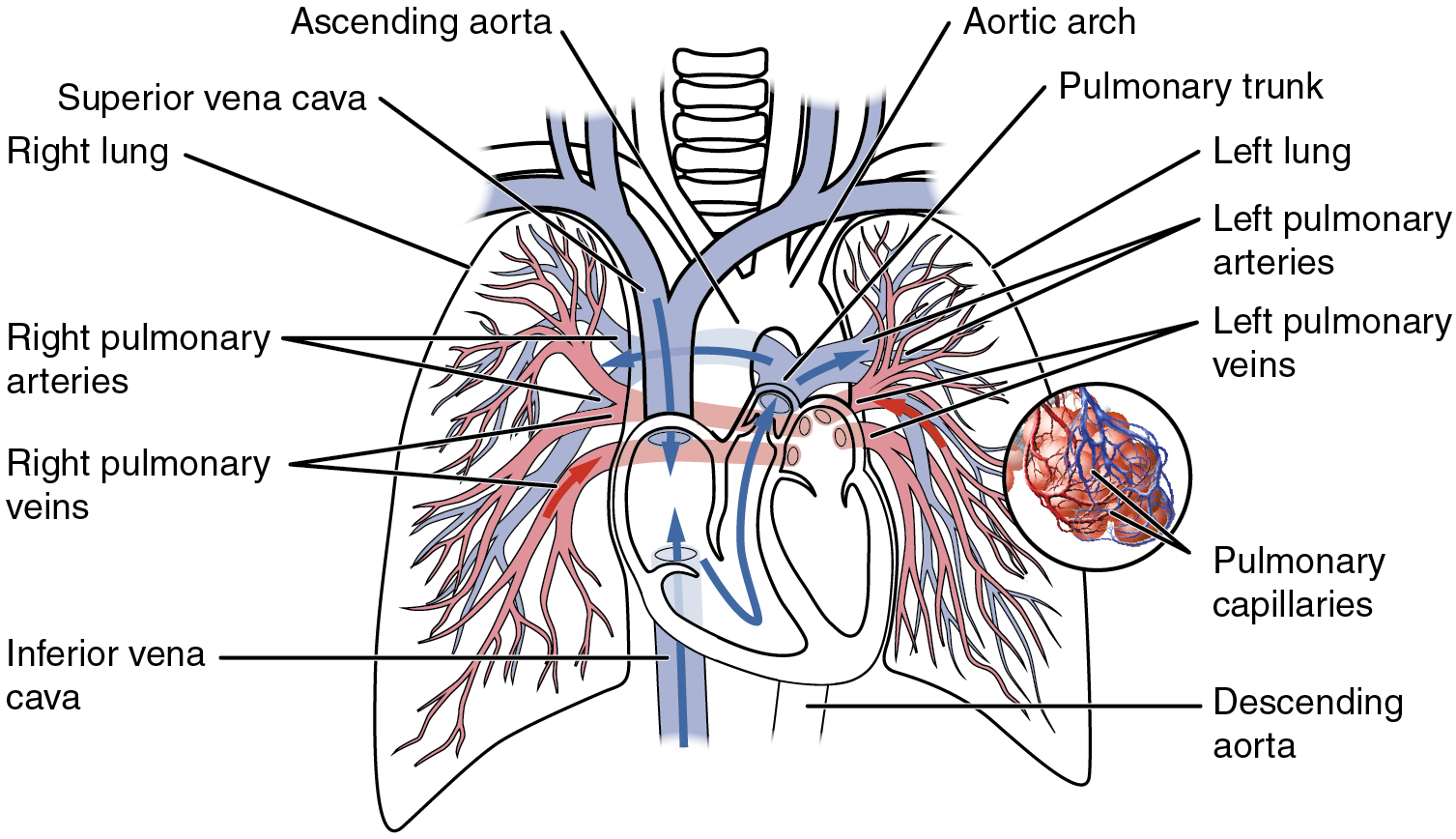
Figure 20.2: The pulmonary circulation as it passes from the heart. Showing both the pulmonary and bronchial arteries.
The pulmonary circulation forms a“loop” through the lungs where blood is oxygenated; and the systemic circulation forms a “loop” through the rest of the body to provide oxygenated blood. The systemic circulation can also be seen to function in two parts – a macrocirculation and a microcirculation. An average adult contains five to six quarts (roughly 4.7 to 5.7 liters) of blood, accounting for approximately 7% of their total body weight. Blood consists of plasma, red blood cells, white blood cells, and platelets. Also, the digestive system works with the circulatory system to provide the nutrients the system needs to keep the heart pumping.
The cardiovascular system of humans is closed, meaning that the blood never leaves the network of blood vessels. In contrast, oxygen and nutrients diffuse across the blood vessel layers and enter interstitial fluid, which carries oxygen and nutrients to the target cells, and carbon dioxide and wastes in the opposite direction.
20.5 The Heart
In humans, the heart is approximately the size of a closed fist and is located between the lungs, in the middle compartment of the chest.
In humans, other mammals, and birds, the heart is divided into four chambers: upper left and right atria and lower left and right ventricles. Commonly the right atrium and ventricle are referred together as the right heart and their left counterparts as the left heart. Fish, in contrast, have two chambers, an atrium and a ventricle, while reptiles have three chambers. In a healthy heart blood flows one way through the heart due to heart valves, which prevent backflow. The heart is enclosed in a protective sac, the pericardium, which also contains a small amount of fluid. The wall of the heart is made up of three layers: epicardium, myocardium, and endocardium.
The heart pumps blood with a rhythm determined by a group of pacemaking cells in the sinoatrial node. These generate a current that causes contraction of the heart, traveling through the atrioventricular node and along the conduction system of the heart. The heart receives blood low in oxygen from the systemic circulation, which enters the right atrium from the superior and inferior venae cavae and passes to the right ventricle. From here it is pumped into the pulmonary circulation, through the lungs where it receives oxygen and gives off carbon dioxide. Oxygenated blood then returns to the left atrium, passes through the left ventricle and is pumped out through the aorta to the systemic circulation−where the oxygen is used and metabolized to carbon dioxide. The heart beats at a resting rate close to 72 beats per minute. Exercise temporarily increases the rate, but lowers resting heart rate in the long term, and is good for heart health.
Cardiovascular diseases (CVD) are the most common cause of death globally as of 2008, accounting for 30% of deaths. Of these more than three quarters are a result of coronary artery disease and stroke. Risk factors include: smoking, being overweight, little exercise, high cholesterol, high blood pressure, and poorly controlled diabetes, among others. Cardiovascular diseases frequently do not have symptoms or may cause chest pain or shortness of breath. Diagnosis of heart disease is often done by the taking of a medical history, listening to the heart-sounds with a stethoscope, ECG, and ultrasound. Specialists who focus on diseases of the heart are called cardiologists, although many specialties of medicine may be involved in treatment.
The human heart is situated in the middle mediastinum, at the level of thoracic vertebrae T5-T8. A double-membraned sac called the pericardium surrounds the heart and attaches to the mediastinum. The back surface of the heart lies near the vertebral column, and the front surface sits behind the sternum and rib cartilages. The upper part of the heart is the attachment point for several large blood vessels—the venae cavae, aorta and pulmonary trunk. The upper part of the heart is located at the level of the third costal cartilage. The lower tip of the heart, the apex, lies to the left of the sternum (8 to 9 cm from the midsternal line) between the junction of the fourth and fifth ribs near their articulation with the costal cartilages.
The largest part of the heart is usually slightly offset to the left side of the chest (though occasionally it may be offset to the right) and is felt to be on the left because the left heart is stronger and larger, since it pumps to all body parts. Because the heart is between the lungs, the left lung is smaller than the right lung and has a cardiac notch in its border to accommodate the heart. The heart is cone-shaped, with its base positioned upwards and tapering down to the apex. An adult heart has a mass of 250–350 grams (9–12 oz). The heart is often described as the size of a fist: 12 cm (5 in) in length, 8 cm (3.5 in) wide, and 6 cm (2.5 in) in thickness, although this description is disputed, as the heart is likely to be slightly larger. Well-trained athletes can have much larger hearts due to the effects of exercise on the heart muscle, similar to the response of skeletal muscle.
20.5.1 Structure of The Heart
The heart has four chambers, two upper atria, the receiving chambers, and two lower ventricles, the discharging chambers. The atria open into the ventricles via the atrioventricular valves, present in the atrioventricular septum. This distinction is visible also on the surface of the heart as the coronary sulcus. There is an ear-shaped structure in the upper right atrium called the right atrial appendage, or auricle, and another in the upper left atrium, the left atrial appendage. The right atrium and the right ventricle together are sometimes referred to as the right heart. Similarly, the left atrium and the left ventricle together are sometimes referred to as the left heart. The ventricles are separated from each other by the interventricular septum, visible on the surface of the heart as the anterior longitudinal sulcus and the posterior interventricular sulcus.
.svg)
Figure 20.3: Diagram of the human heart 1. Superior vena cava 2. 4. Mitral valve 5. Aortic valve 6. Left ventricle 7. Right ventricle 8. Left atrium 9. Right atrium 10. Aorta 11. Pulmonary valve 12. Tricuspid valve 13. Inferior vena cava
The cardiac skeleton is made of dense connective tissue and this gives structure to the heart. It forms the atrioventricular septum which separates the atria from the ventricles, and the fibrous rings which serve as bases for the four heart valves. The cardiac skeleton also provides an important boundary in the heart’s electrical conduction system since collagen cannot conduct electricity. The interatrial septum separates the atria and the interventricular septum separates the ventricles. The interventricular septum is much thicker than the interatrial septum, since the ventricles need to generate greater pressure when they contract.
The heart has four valves, which separate its chambers. One valve lies between each atrium and ventricle, and one valve rests at the exit of each ventricle.
The valves between the atria and ventricles are called the atrioventricular valves. Between the right atrium and the right ventricle is the tricuspid valve. The tricuspid valve has three cusps, which connect to chordae tendinae and three papillary muscles named the anterior, posterior, and septal muscles, after their relative positions. The mitral valve lies between the left atrium and left ventricle. It is also known as the bicuspid valve due to its having two cusps, an anterior and a posterior cusp. These cusps are also attached via chordae tendinae to two papillary muscles projecting from the ventricular wall.
The papillary muscles extend from the walls of the heart to valves by cartilaginous connections called chordae tendinae. These muscles prevent the valves from falling too far back when they close. During the relaxation phase of the cardiac cycle, the papillary muscles are also relaxed and the tension on the chordae tendineae is slight. As the heart chambers contract, so do the papillary muscles. This creates tension on the chordae tendineae, helping to hold the cusps of the atrioventricular valves in place and preventing them from being blown back into the atria. [g]
Two additional semilunar valves sit at the exit of each of the ventricles. The pulmonary valve is located at the base of the pulmonary artery. This has three cusps which are not attached to any papillary muscles. When the ventricle relaxes blood flows back into the ventricle from the artery and this flow of blood fills the pocket-like valve, pressing against the cusps which close to seal the valve. The semilunar aortic valve is at the base of the aorta and also is not attached to papillary muscles. This too has three cusps which close with the pressure of the blood flowing back from the aorta.
The right heart consists of two chambers, the right atrium and the right ventricle, separated by a valve, the tricuspid valve.
The right atrium receives blood almost continuously from the body’s two major veins, the superior and inferior venae cavae. A small amount of blood from the coronary circulation also drains into the right atrium via the coronary sinus, which is immediately above and to the middle of the opening of the inferior vena cava. In the wall of the right atrium is an oval-shaped depression known as the fossa ovalis, which is a remnant of an opening in the fetal heart known as the foramen ovale. Most of the internal surface of the right atrium is smooth, the depression of the fossa ovalis is medial, and the anterior surface has prominent ridges of pectinate muscles, which are also present in the right atrial appendage.
The right atrium is connected to the right ventricle by the tricuspid valve. The walls of the right ventricle are lined with trabeculae carneae, ridges of cardiac muscle covered by endocardium. In addition to these muscular ridges, a band of cardiac muscle, also covered by endocardium, known as the moderator band reinforces the thin walls of the right ventricle and plays a crucial role in cardiac conduction. It arises from the lower part of the interventricular septum and crosses the interior space of the right ventricle to connect with the inferior papillary muscle. The right ventricle tapers into the pulmonary trunk, into which it ejects blood when contracting. The pulmonary trunk branches into the left and right pulmonary arteries that carry the blood to each lung. The pulmonary valve lies between the right heart and the pulmonary trunk.
The left heart has two chambers: the left atrium and the left ventricle, separated by the mitral valve.
The left atrium receives oxygenated blood back from the lungs via one of the four pulmonary veins. The left atrium has an outpouching called the left atrial appendage. Like the right atrium, the left atrium is lined by pectinate muscles. The left atrium is connected to the left ventricle by the mitral valve.
The left ventricle is much thicker as compared with the right, due to the greater force needed to pump blood to the entire body. Like the right ventricle, the left also has trabeculae carneae, but there is no moderator band. The left ventricle pumps blood to the body through the aortic valve and into the aorta. Two small openings above the aortic valve carry blood to the heart itself, the left main coronary artery and the right coronary artery.
The heart wall is made up of three layers: the inner endocardium, middle myocardium and outer epicardium. These are surrounded by a double-membraned sac called the pericardium.
The innermost layer of the heart is called the endocardium. It is made up of a lining of simple squamous epithelium and covers heart chambers and valves. It is continuous with the endothelium of the veins and arteries of the heart, and is joined to the myocardium with a thin layer of connective tissue. The endocardium, by secreting endothelins, may also play a role in regulating the contraction of the myocardium.
The middle layer of the heart wall is the myocardium, which is the cardiac muscle—a layer of involuntary striated muscle tissue surrounded by a framework of collagen. The cardiac muscle pattern is elegant and complex, as the muscle cells swirl and spiral around the chambers of the heart, with the outer muscles forming a figure 8 pattern around the atria and around the bases of the great vessels and the inner muscles, forming a figure 8 around the two ventricles and proceeding toward the apex. This complex swirling pattern allows the heart to pump blood more effectively.
There are two types of cells in cardiac muscle: muscle cells which have the ability to contract easily, and pacemaker cells of the conducting system. The muscle cells make up the bulk (99%) of cells in the atria and ventricles. These contractile cells are connected by intercalated discs which allow a rapid response to impulses of action potential from the pacemaker cells. The intercalated discs allow the cells to act as a syncytium and enable the contractions that pump blood through the heart and into the major arteries. The pacemaker cells make up 1% of cells and form the conduction system of the heart. They are generally much smaller than the contractile cells and have few myofibrils which gives them limited contractibility. Their function is similar in many respects to neurons. Cardiac muscle tissue has autorhythmicity, the unique ability to initiate a cardiac action potential at a fixed rate—spreading the impulse rapidly from cell to cell to trigger the contraction of the entire heart.
20.5.2 The Pericardium
The pericardium is the sack that surrounds the heart. The tough outer surface of the pericardium is called the fibrous membrane. This is lined by a double inner membrane called the serous membrane that produces pericardial fluid to lubricate the surface of the heart. The part of the serous membrane attached to the fibrous membrane is called the parietal pericardium, while the part of the serous membrane attached to the heart is known as the visceral pericardium. The pericardium is present in order to lubricate its movement against other structures within the chest, to keep the heart’s position stabilised within the chest, and to protect the heart from infection.
Heart tissue, like all cells in the body, needs to be supplied with oxygen, nutrients and a way of removing metabolic wastes. This is achieved by the coronary circulation, which includes arteries, veins, and lymphatic vessels. Blood flow through the coronary vessels occurs in peaks and troughs relating to the heart muscle’s relaxation or contraction.
Heart tissue receives blood from two arteries which arise just above the aortic valve. These are the left main coronary artery and the right coronary artery. The left main coronary artery splits shortly after leaving the aorta into two vessels, the left anterior descending and the left circumflex artery. The left anterior descending artery supplies heart tissue and the front, outer side, and the septum of the left ventricle. It does this by branching into smaller arteries—diagonal and septal branches. The left circumflex supplies the back and underneath of the left ventricle. The right coronary artery supplies the right atrium, right ventricle, and lower posterior sections of the left ventricle. The right coronary artery also supplies blood to the atrioventricular node (in about 90% of people) and the sinoatrial node (in about 60% of people). The right coronary artery runs in a groove at the back of the heart and the left anterior descending artery runs in a groove at the front. There is significant variation between people in the anatomy of the arteries that supply the heart The arteries divide at their furtherst reaches into smaller branches that join together at the edges of each arterial distribution.

The coronary sinus is a large vein that drains into the right atrium, and receives most of the venous drainage of the heart. It receives blood from the great cardiac vein (receiving the left atrium and both ventricles), the posterior cardiac vein (draining the back of the left ventricle), the middle cardiac vein (draining the bottom of the left and right ventricles), and small cardiac veins. The anterior cardiac veins drain the front of the right ventricle and drain directly into the right atrium.
The heart receives nerve signals from the vagus nerve and from nerves arising from the sympathetic trunk. These nerves act to influence, but not control, the heart rate. Sympathetic nerves also influence the force of heart contraction. Signals that travel along these nerves arise from two paired cardiovascular centres in the medulla oblongata. The vagus nerve of the parasympathetic nervous system acts to decrease the heart rate, and nerves from the sympathetic trunk act to increase the heart rate. These nerves form a network of nerves that lies over the heart called the cardiac plexus.
The vagus nerve is a long, wandering nerve that emerges from the brainstem and provides parasympathetic stimulation to a large number of organs in the thorax and abdomen, including the heart. The nerves from the sympathetic trunk emerge through the T1-T4 thoracic ganglia and travel to both the sinoatrial and atrioventricular nodes, as well as to the atria and ventricles. The ventricles are more richly innervated by sympathetic fibers than parasympathetic fibers. Sympathetic stimulation causes the release of the neurotransmitter norepinephrine (also known as noradrenaline) at the neuromuscular junction of the cardiac nerves. This shortens the repolarization period, thus speeding the rate of depolarization and contraction, which results in an increased heart rate. It opens chemical or ligand-gated sodium and calcium ion channels, allowing an influx of positively charged ions. Norepinephrine binds to the beta–1 receptor.
20.6 The Blood Flow
The heart functions as a pump in the circulatory system to provide a continuous flow of blood throughout the body. This circulation consists of the systemic circulation to and from the body and the pulmonary circulation to and from the lungs. Blood in the pulmonary circulation exchanges carbon dioxide for oxygen in the lungs through the process of respiration. The systemic circulation then transports oxygen to the body and returns carbon dioxide and relatively deoxygenated blood to the heart for transfer to the lungs.
The right heart collects deoxygenated blood from two large veins, the superior and inferior venae cavae. Blood collects in the right and left atrium continuously. The superior vena cava drains blood from above the diaphragm and empties into the upper back part of the right atrium. The inferior vena cava drains the blood from below the diaphragm and empties into the back part of the atrium below the opening for the superior vena cava. Immediately above and to the middle of the opening of the inferior vena cava is the opening of the thin-walled coronary sinus. Additionally, the coronary sinus returns deoxygenated blood from the myocardium to the right atrium. The blood collects in the right atrium. When the right atrium contracts, the blood is pumped through the tricuspid valve into the right ventricle. As the right ventricle contracts, the tricuspid valve closes and the blood is pumped into the pulmonary trunk through the pulmonary valve. The pulmonary trunk divides into pulmonary arteries and progressively smaller arteries throughout the lungs, until it reaches capillaries. As these pass by alveoli carbon dioxide is exchanged for oxygen. This happens through the passive process of diffusion.
In the left heart, oxygenated blood is returned to the left atrium via the pulmonary veins. It is then pumped into the left ventricle through the mitral valve and into the aorta through the aortic valve for systemic circulation. The aorta is a large artery that branches into many smaller arteries, arterioles, and ultimately capillaries. In the capillaries, oxygen and nutrients from blood are supplied to body cells for metabolism, and exchanged for carbon dioxide and waste products. Capillary blood, now deoxygenated, travels into venules and veins that ultimately collect in the superior and inferior vena cavae, and into the right heart.
20.6.1 The Cardiac Cycle
The cardiac cycle refers to the sequence of events in which the heart contracts and relaxes with every heartbeat. The period of time during which the ventricles contract, forcing blood out into the aorta and main pulmonary artery, is known as systole, while the period during which the ventricles relax and refill with blood is known as diastole. The atria and ventricles work in concert, so in systole when the ventricles are contracting, the atria are relaxed and collecting blood. When the ventricles are relaxed in diastole, the atria contract to pump blood to the ventricles. This coordination ensures blood is pumped efficiently to the body.
At the beginning of the cardiac cycle, the ventricles are relaxing. As they do so, they are filled by blood passing through the open mitral and tricuspid valves. After the ventricles have completed most of their filling, the atria contract, forcing further blood into the ventricles and priming the pump. Next, the ventricles start to contract. As the pressure rises within the cavities of the ventricles, the mitral and tricuspid valves are forced shut. As the pressure within the ventricles rises further, exceeding the pressure with the aorta and pulmonary arteries, the aortic and pulmonary valves open. Blood is ejected from the heart, causing the pressure within the ventricles to fall. Simultaneously, the atria refill as blood flows into the right atrium through the superior and inferior vena cavae, and into the left atrium through the pulmonary veins. Finally, when the pressure within the ventricles falls below the pressure within the aorta and pulmonary arteries, the aortic and pulmonary valves close. The ventricles start to relax, the mitral and tricuspid valves open, and the cycle begins again
20.6.2 Control of The Heart Rate
The normal resting heart rate is called the sinus rhythm, created and sustained by the sinoatrial node, a group of pacemaking cells found in the wall of the right atrium. Cells in the sinoatrial node do this by creating an action potential. The cardiac action potential is created by the movement of specific electrolytes into and out of the pacemaker cells. The action potential then spreads to nearby cells.

Figure 20.5: Conduction system of the heart.
When the sinoatrial cells are resting, they have a negative charge on their membranes. However a rapid influx of sodium ions causes the membrane’s charge to become positive. This is called depolarisation and occurs spontaneously. Once the cell has a sufficiently high charge, the sodium channels close and calcium ions then begin to enter the cell, shortly after which potassium begins to leave it. All the ions travel through ion channels in the membrane of the sinoatrial cells. The potassium and calcium start to move out of and into the cell only once it has a sufficiently high charge, and so are called voltage-gated. Shortly after this, the calcium channels close and potassium channels open, allowing potassium to leave the cell. This causes the cell to have a negative resting charge and is called repolarization. When the membrane potential reaches approximately −60 mV, the potassium channels close and the process may begin again.
The ions move from areas where they are concentrated to where they are not. For this reason sodium moves into the cell from outside, and potassium moves from within the cell to outside the cell. Calcium also plays a critical role. Their influx through slow channels means that the sinoatrial cells have a prolonged “plateau” phase when they have a positive charge. A part of this is called the absolute refractory period. Calcium ions also combine with the regulatory protein troponin C in the troponin complex to enable contraction of the cardiac muscle, and separate from the protein to allow relaxation.
The adult resting heart rate ranges from 60 to 100 bpm. The resting heart rate of a newborn can be 129 beats per minute (bpm) and this gradually decreases until maturity. An athlete’s heart rate can be lower than 60 bpm. During exercise the rate can be 150 bpm with maximum rates reaching from 200 to 220 bpm.
20.6.3 The Electrocardiogram (EKG)
Using surface electrodes on the body, it is possible to record the electrical activity of the heart. This tracing of the electrical signal is the electrocardiogram (ECG) or (EKG). An ECG is a bedside test and involves the placement of ten leads on the body. This produces a “12 lead” ECG (three extra leads are calculated mathematically, and one lead is a ground).
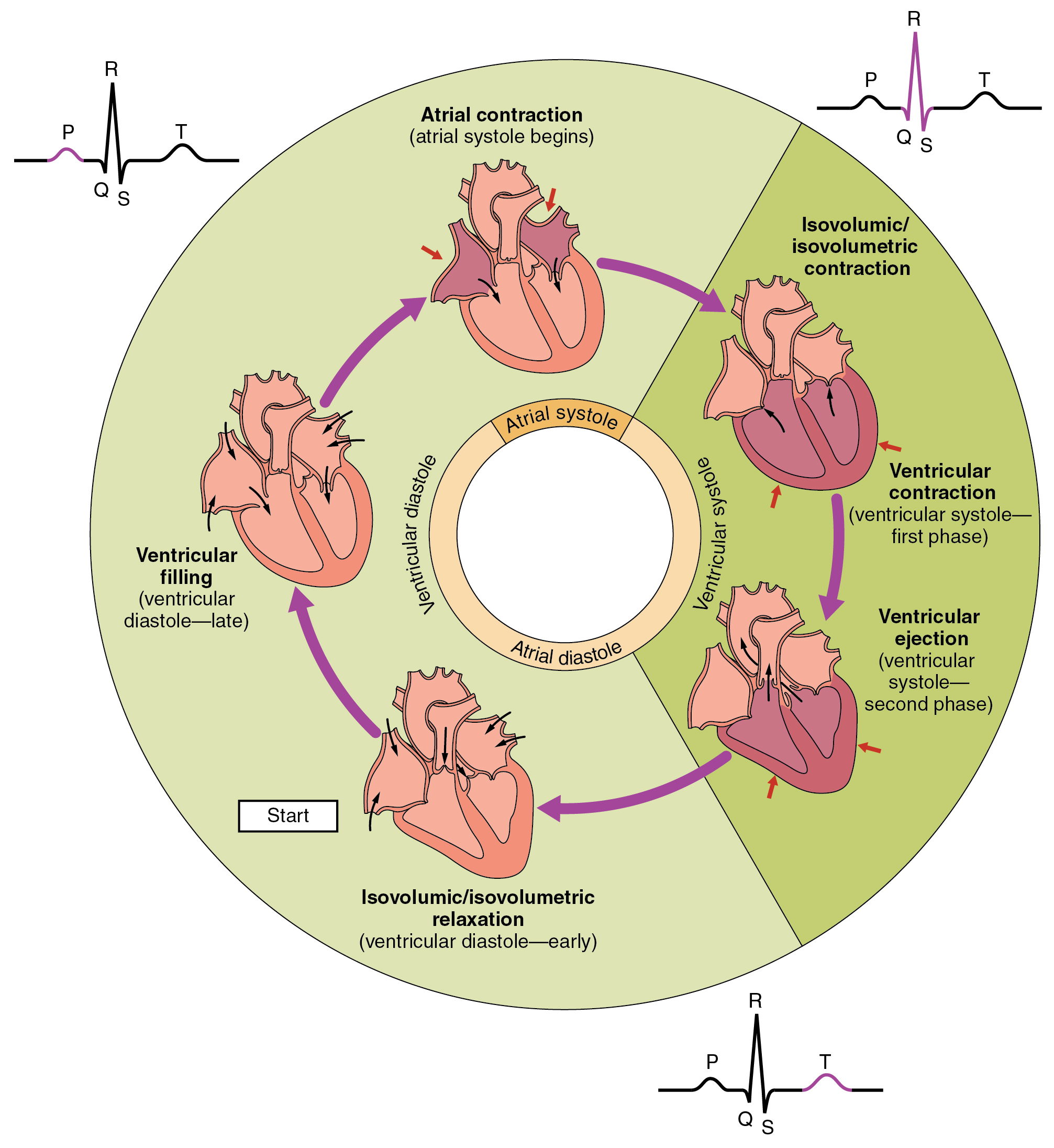
Figure 20.6: The cardiac cycle as correlated to the EKG
There are five prominent features on the ECG: the P wave (atrial depolarisation), the QRS complex (ventricular depolarisation) and the T wave (ventricular repolarisation). As the heart cells contract, they create a current that travels through the heart. A downward deflection on the ECG implies cells are becoming more positive in charge (“depolarising”) in the direction of that lead, whereas an upward inflection implies cells are becoming more negative (“repolarising”) in the direction of the lead. This depends on the position of the lead, so if a wave of depolarising moved from left to right, a lead on the left would show a negative deflection, and a lead on the right would show a positive deflection. The ECG is a useful tool in detecting rhythm disturbances and in detecting insufficient blood supply to the heart. Sometimes abnormalities are suspected, but not immediately visible on the ECG. Testing when exercising can be used to provoke an abnormality, or an ECG can be worn for a longer period such as a 24-hour Holter monitor if a suspected rhythm abnormality is not present at the time of assessment.
20.7 Arteries
Oxygenated blood enters the systemic circulation when leaving the left ventricle, through the aortic semilunar valve. The first part of the systemic circulation is the aorta, a massive and thick-walled artery. The aorta arches and gives branches supplying the upper part of the body after passing through the aortic opening of the diaphragm at the level of thoracic ten vertebra, it enters the abdomen. Later it descends down and supplies branches to abdomen, pelvis, perineum and the lower limbs. The walls of aorta are elastic. This elasticity helps to maintain the blood pressure throughout the body. When the aorta receives almost five litres of blood from the heart, it recoils and is responsible for pulsating blood pressure. Moreover, as aorta branches into smaller arteries, their elasticity goes on decreasing and their compliance goes on increasing.
20.8 Capillaries
Arteries branch into small passages called arterioles and then into the capillaries. The capillaries merge to bring blood into the venous system.
20.9 Veins
Capillaries merge into venules, which merge into veins. The venous system feeds into the two major veins: the superior vena cava – which mainly drains tissues above the heart – and the inferior vena cava – which mainly drains tissues below the heart. These two large veins empty into the right atrium of the heart.
20.9.1 Portal veins
The general rule is that arteries from the heart branch out into capillaries, which collect into veins leading back to the heart. Portal veins are a slight exception to this. In humans the only significant example is the hepatic portal vein which combines from capillaries around the gastrointestinal tract where the blood absorbs the various products of digestion; rather than leading directly back to the heart, the hepatic portal vein branches into a second capillary system in the liver.
20.10 The Systemic Circulation
Systemic circulation is the portion of the cardiovascular system which transports oxygenated blood away from the heart through the aorta from the left ventricle where the blood has been previously deposited from pulmonary circulation, to the rest of the body, and returns oxygen-depleted blood back to the heart.
The brain has a dual blood supply that comes from arteries at its front and back. These are called the “anterior” and “posterior” circulation respectively. The anterior circulation arises from the internal carotid arteries and supplies the front of the brain. The posterior circulation arises from the vertebral arteries, and supplies the back of the brain and brainstem. The circulation from the front and the back join together (anastomise) at the Circle of Willis.
The renal circulation receives around 20% of the cardiac output. It branches from the abdominal aorta and returns blood to the ascending vena cava. It is the blood supply to the kidneys, and contains many specialized blood vessels.
20.11 The Pulmonary Circulation
The circulatory system of the lungs is the portion of the cardiovascular system in which oxygen-depleted blood is pumped away from the heart, via the pulmonary artery, to the lungs and returned, oxygenated, to the heart via the pulmonary vein.
Oxygen deprived blood from the superior and inferior vena cava enters the right atrium of the heart and flows through the tricuspid valve (right atrioventricular valve) into the right ventricle, from which it is then pumped through the pulmonary semilunar valve into the pulmonary artery to the lungs. Gas exchange occurs in the lungs, whereby CO 2 is released from the blood, and oxygen is absorbed. The pulmonary vein returns the now oxygen-rich blood to the left atrium.
A separate system known as the bronchial circulation supplies blood to the tissue of the larger airways of the lung.
20.12 Cardiovascular Disease
Diseases affecting the cardiovascular system are called cardiovascular disease.
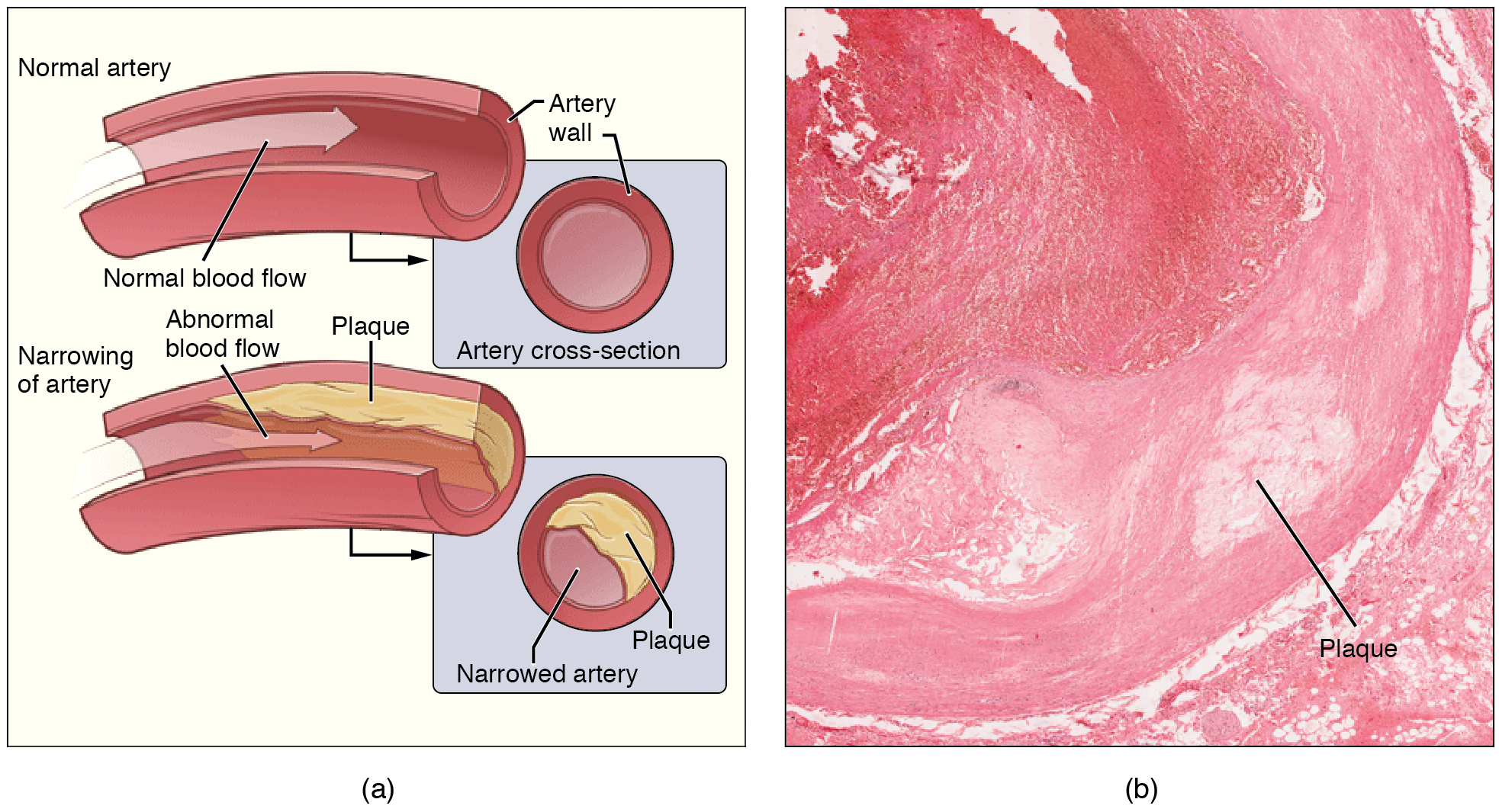
Many of these diseases are called “lifestyle diseases” because they develop over time and are related to a person’s exercise habits, diet, whether they smoke, and other lifestyle choices a person makes. Atherosclerosis is the precursor to many of these diseases. It is where small atheromatous plaques build up in the walls of medium and large arteries. This may eventually grow or rupture to occlude the arteries. It is also a risk factor for acute coronary syndromes, which are diseases that are characterised by a sudden deficit of oxygenated blood to the heart tissue. Atherosclerosis is also associated with problems such as aneurysm formation or splitting (“dissection”) of arteries.
{kind=link}
{kind=link}
.svg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Another major cardiovascular disease involves the creation of a clot, called a “thrombus”. These can originate in veins or arteries. Deep venous thrombosis, which mostly occurs in the legs, is one cause of clots in the veins of the legs, particularly when a person has been stationary for a long time. These clots may embolise, meaning travel to another location in the body. The results of this may include pulmonary embolus, transient ischaemic attacks, or stroke.
Cardiovascular diseases may also be congenital in nature, such as heart defects or persistent fetal circulation, where the circulatory changes that are supposed to happen after birth do not. Not all congenital changes to the circulatory system are associated with diseases, a large number are anatomical variations.
20.13 History
The earliest known writings on the circulatory system are found in the Ebers Papyrus (16th century BCE), an ancient Egyptian medical papyrus containing over 700 prescriptions and remedies, both physical and spiritual. In the papyrus, it acknowledges the connection of the heart to the arteries. The Egyptians thought air came in through the mouth and into the lungs and heart. From the heart, the air travelled to every member through the arteries. Although this concept of the circulatory system is only partially correct, it represents one of the earliest accounts of scientific thought.
In the 6th century BCE, the knowledge of circulation of vital fluids through the body was known to the Ayurvedic physician Sushruta in ancient India. He also seems to have possessed knowledge of the arteries, described as ‘channels’ by Dwivedi & Dwivedi (2007). The valves of the heart were discovered by a physician of the Hippocratean school around the 4th century BCE. However their function was not properly understood then. Because blood pools in the veins after death, arteries look empty. Ancient anatomists assumed they were filled with air and that they were for transport of air.
The Greek physician, Herophilus, distinguished veins from arteries but thought that the pulse was a property of arteries themselves. Greek anatomist Erasistratus observed that arteries that were cut during life bleed. He ascribed the fact to the phenomenon that air escaping from an artery is replaced with blood that entered by very small vessels between veins and arteries. Thus he apparently postulated capillaries but with reversed flow of blood.
In 2nd century AD Rome, the Greek physician Galen knew that blood vessels carried blood and identified venous (dark red) and arterial (brighter and thinner) blood, each with distinct and separate functions. Growth and energy were derived from venous blood created in the liver from chyle, while arterial blood gave vitality by containing pneuma (air) and originated in the heart. Blood flowed from both creating organs to all parts of the body where it was consumed and there was no return of blood to the heart or liver. The heart did not pump blood around, the heart’s motion sucked blood in during diastole and the blood moved by the pulsation of the arteries themselves.
Galen believed that the arterial blood was created by venous blood passing from the left ventricle to the right by passing through ‘pores’ in the interventricular septum, air passed from the lungs via the pulmonary artery to the left side of the heart. As the arterial blood was created ‘sooty’ vapors were created and passed to the lungs also via the pulmonary artery to be exhaled.
In 1025, The Canon of Medicine by the Persian physician, Avicenna, “erroneously accepted the Greek notion regarding the existence of a hole in the ventricular septum by which the blood traveled between the ventricles.” Despite this, Avicenna “correctly wrote on the cardiac cycles and valvular function”, and “had a vision of blood circulation” in his Treatise on Pulse. While also refining Galen’s erroneous theory of the pulse, Avicenna provided the first correct explanation of pulsation: “Every beat of the pulse comprises two movements and two pauses. Thus, expansion : pause : contraction : pause. […] The pulse is a movement in the heart and arteries … which takes the form of alternate expansion and contraction.”
In 1242, the Arabian physician, Ibn al-Nafis, became the first person to accurately describe the process of pulmonary circulation, for which he is sometimes considered the father of circulatory physiology.[failed verification] Ibn al-Nafis stated in his Commentary on Anatomy in Avicenna’s Canon:
“…the blood from the right chamber of the heart must arrive at the left chamber but there is no direct pathway between them. The thick septum of the heart is not perforated and does not have visible pores as some people thought or invisible pores as Galen thought. The blood from the right chamber must flow through the vena arteriosa (pulmonary artery) to the lungs, spread through its substances, be mingled there with air, pass through the arteria venosa (pulmonary vein) to reach the left chamber of the heart and there form the vital spirit…”
In addition, Ibn al-Nafis had an insight into what would become a larger theory of the capillary circulation. He stated that “there must be small communications or pores (manafidh in Arabic) between the pulmonary artery and vein,” a prediction that preceded the discovery of the capillary system by more than 400 years. Ibn al-Nafis’ theory, however, was confined to blood transit in the lungs and did not extend to the entire body.
Michael Servetus was the first European to describe the function of pulmonary circulation, although his achievement was not widely recognized at the time, for a few reasons. He firstly described it in the “Manuscript of Paris” (near 1546), but this work was never published. And later he published this description, but in a theological treatise, Christianismi Restitutio, not in a book on medicine. Only three copies of the book survived but these remained hidden for decades, the rest were burned shortly after its publication in 1553 because of persecution of Servetus by religious authorities.
Better known discovery of pulmonary circulation was by Vesalius’s successor at Padua, Realdo Colombo, in 1559.
Finally, the English physician William Harvey, a pupil of Hieronymus Fabricius (who had earlier described the valves of the veins without recognizing their function), performed a sequence of experiments and published his Exercitatio Anatomica de Motu Cordis et Sanguinis in Animalibus in 1628, which “demonstrated that there had to be a direct connection between the venous and arterial systems throughout the body, and not just the lungs. Most importantly, he argued that the beat of the heart produced a continuous circulation of blood through minute connections at the extremities of the body. This is a conceptual leap that was quite different from Ibn al-Nafis’ refinement of the anatomy and bloodflow in the heart and lungs.” This work, with its essentially correct exposition, slowly convinced the medical world. However, Harvey was not able to identify the capillary system connecting arteries and veins; these were later discovered by Marcello Malpighi in 1661.
In 1956, André Frédéric Cournand, Werner Forssmann and Dickinson W. Richards were awarded the Nobel Prize in Medicine “for their discoveries concerning heart catheterization and pathological changes in the circulatory system.” In his Nobel lecture, Forssmann credits Harvey as birthing cardiology with the publication of his book in 1628.
In the 1970s, Diana McSherry developed computer-based systems to create images of the circulatory system and heart without the need for surgery.